

As per the request in this thread, I am starting this thread to debate the question of common descent. I'm perfectly happy to kick off the discussion with a presentation of a tiny slice of the evidence biologists consider established common descent.
Common descent is a historical claim about the evolutionary past of organisms. It refers to the idea that all organisms are related to one another like a gigantic extended family. For example: five million years ago, chimpanzees and humans did not exist separately, but were represented by a single species. Approximately 4.5 million years ago that population underwent speciation and each new species (at that point very similar to one another), gradually diverged. According to all of mainstream biology, this principle holds true for all life, so that every species shares ancestry with every other at some point in the distant past.
To make this claim, biologists have a large amount of evidence that they think establishes common descent as a very robust scientific claim. The famous talkorigins '20+ evidences for macroevolution' article is actually a list of evidence for common descent, and doesn't actually have much to do with macroevolution as modern biology uses the term.
Rather than use that article here, I will instead go strait to the most authoritative sources I have access to, and give a quick summary of the overview of the evidence from Douglas J. Futuymas Evolutionary Biology 3rd edition, the most respected general textbook in the field.
Futuyma lists eight categories of evidence, each of which would require a fair amount of elaboration. I'll give the full list before expanding on one or two.
1. The hierarchical organisation of life.
2. Homology
3. Embryological similarities
4. Vestigial characters
5. Convergence
6. Suboptimal design
7. Geographical distributions
and 8. Intermediate forms.
I'll expand a little on 1 and 8 before throwing open the floor.
Life on earth exists in a nested hierarchy. This means that if you divide organisms into groups according to their features, you can then divide each of those groups into more groups without overlapping. So, if one divides the tetrapods from the arthropods, one can then pick out smaller groups; say, the mammals out of the tetrapods, and the insects from the arthropods, and those groups will not overlap in any new features that are not present in their parent groups. Meanwhile, if you try to group cars first by engine size, and then by colour, you'll find that there is a lot of overlap of features in groups that aren't present in those groups parent categories. So both red cars and blue cars can have both V6 and V12 engines, but triple segmentation with six legs is found only in arthropod groups (namely insects), while milk is found only in tetrapod groups (namely mammals).
This is evidence for common descent, because this pattern of nested hierarchy is produced by a historic pattern of descent and divergence, and by hardly any other types of processes. For example, tracing the surnames in a family tree yields a nested hierarchy: if one branch of the plantaginet family changes its name to plantaginet-smith, then you'll find plantaginet smith only in the larger plantaginet extended family. Only objects that have been produced by descent and branching fall into true nested hierarchies.
This fact about living things allows us to do all sorts of things in the exciting world of molecular biology. Nested hierarchies are what enable us to produce phylogenetic trees of life, which are essentially hypotheses about a particular pattern of descent. Using the same methods we now use to establish paternity from blood samples in court, we establish the relationships of species. The most striking thing is that we can yield the same phylogenetic tree by analysing just about any genetic feature: be it a gene, or mitochondrial DNA, or a particular protein sequence like hemoglobin, or most tellingly, endogenous retroviral insertions (I can expand on this last one if it's unfamiliar), the same tree keeps appearing again and again. This would be inexplicable if all species had separate origins, but is exactly what common descent predicts.
Now, to intermediate forms. Here, I will list the fossilised forms of three major evolutionary lineages, from three of my favourite books. I can't supply the diagrams, obviously, but I will give page numbers for anyone who wants to check up on me.
First, it should be established that according to the scenario of independent creation of organisms, intermediate sequences should not exist at all. Given that, the following three gradual evolutionary sequences establish the evolutionary history of these organisms: lizards to mammals, four-legged artiodoctyls to whales, and hyracotherium to modern horses.
From Ridleys Evolution text: The following fossils form a transitional lineage: Reptilian pelycosaurs to Ophlacodontids (which also gave rise to the famous Dimetrodon ), to the broadly varying Therapsids, from amongst whos number came the Cynodots, representing whom there are many individual specimens, such as(in approximate evolutionary order):
Procynosuchus
Thrinaxodon (here's another thrinaxodon)
Diademodon
Probainognathus
These specimens become increasingly less reptilian and begin to accumulate mammilian features over time. The latest cynodonts in this series blend neatly into the early mammals.
I'm running short on time, so I'll finish whales and horses quickly.
Refer to At the Waters Edge by Carl Zimmer for more information on this graduation from four legged pakicetus to modern whales:
Pakicetus, Ambulocetus, Dalanistes, Rodhocetus, Takracetus, Gaviocetus, Basilisaurus, Dorudon, Mysticetes (modern baleen whales). Googling these names might yield more information as well.
Refer to Strickberger and Monroes' Evolution textbook for more information on the transition from the small 55 million year old Hyracotherium, through the Oligocene Miohippus and Miocene Merychippus to modern Equus. This is a famously well documented transition, so it should be easy to find online resources.
But right now, I've made myself late. I suppose the topic of this debate should be: 'Do these observations, which are commonly advanced by mainstream biology, provide evidence for common descent?'
Good night and enjoy the debate.
The reality or otherwise of Common Descent
Moderator: Moderators
-
otseng

- Savant
- Posts: 20518
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #2
Let me start off by saying I have no subject expertise in the field of biology, so any criticisms of my lack of knowledge in this field would be completely founded. On the other hand, I have no subject expertise in any of the subjects here in this forum and it hasn't prevented me yet in engaging in debating them. 
OK, let's start by discussing the first point that you brought up.
If the mechanism for all evolutionary processes is microevolution (would that be a fair statement?), then why do we have a hierarchical tree? Why do we not see a smooth transition between all life forms (particularly in the fossil record)?
Is it true that all living organisms in the lower part of the tree no longer exist?
By what criteria are living things categorized into the phylogenetic tree?
How exactly does the fact that life can be organized into a hierarchy show that there was common descent? I can classify all the books in my bookshelf into a hierarchy, but it doesn't mean there was a common descent.
OK, let's start by discussing the first point that you brought up.
1. The hierarchical organisation of life.
If the mechanism for all evolutionary processes is microevolution (would that be a fair statement?), then why do we have a hierarchical tree? Why do we not see a smooth transition between all life forms (particularly in the fossil record)?
Is it true that all living organisms in the lower part of the tree no longer exist?
By what criteria are living things categorized into the phylogenetic tree?
How exactly does the fact that life can be organized into a hierarchy show that there was common descent? I can classify all the books in my bookshelf into a hierarchy, but it doesn't mean there was a common descent.
Post #3
I'm not sure what you mean by this. The classification of species into a family tree doesn't by itself say anything about the rate of change that we should expect from the fossil record.otseng wrote: If the mechanism for all evolutionary processes is microevolution (would that be a fair statement?), then why do we have a hierarchical tree? Why do we not see a smooth transition between all life forms (particularly in the fossil record)?
The reason we have a pattern of descent that looks like a bifurcating tree is because each species generally stays homogenous. That's not to say the species doesn't evolve, but it's the nature of mutation/natural selection that the whole of a given interbreeding population will inherit changes in one individual. Given this, when speciation occurs it divides one species into two, which can never again recombine. Thus, the pattern of descent proceeds by bifurcations, as changes in the two new species no longer match one another, and the populations drift apart.
This says nothing about how smooth we expect transitions to be, except that we do not expect extant (currently living) species to grade smoothly in to one another. Instead, biodiversity should look like groups within larger groups withing still larger groups, which is the nested hierarchy we do in fact find.
What we expect from the fossil record depend entirely on geology, evolutionary rates of change, and being downright lucky enough to chance apon the best fossils (who knows how many nice intermediate series we'd have if we could somehow extract every fossil). This means that not having a good intermediate series of fossils could be simple bad luck. Of course, we do have many very good gradual sequences, and I outlined three of them above.
All organisms in the lower part of the tree are dead, but not necessarily extinct. Remember that the phylogenetic tree is very like a family tree: everyone at the top of the tree is a modern person, while people further down are ancestors. Just because a species can be found in the lower branches of the tree doesn't mean you can't still find them alive today: all this means is that there is a branch extending from the ancient species all the way to the top. In the family tree example, you can find plantaginets a good way down the english royal family tree, but that does not mean there are no living plantaginets today. Similarly, there are coelocanth fossils placing the start of the coelocanth branch a fair way down the tree, but the lineage still survives today.Is it true that all living organisms in the lower part of the tree no longer exist?
I hope I'm not being too unclear?
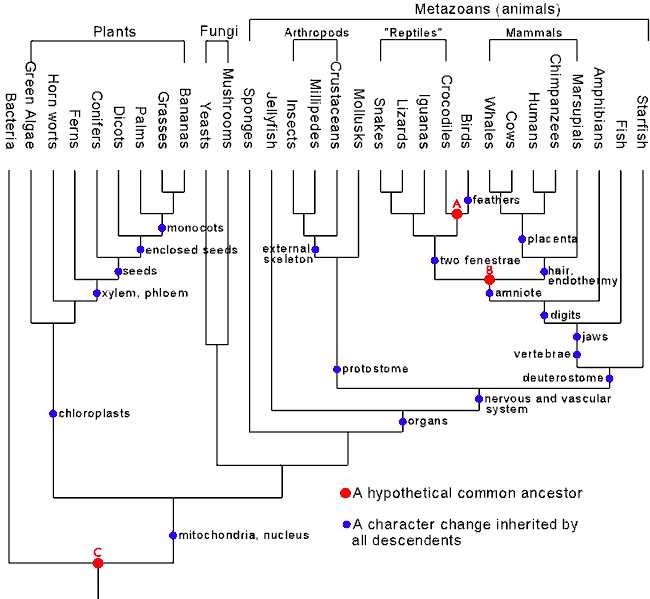
Phylogenetic trees are the realm of an entire field of biology called systematics. Because biodiversity falls into a nested hierarchy, we can build cladograms of chunks of the tree of life. It's important to note, and I'll come back to this, that this would be impossible if life did not share ancestryBy what criteria are living things categorized into the phylogenetic tree?
This is an explanatory diagram from Campbell and Reece Biology 6th edition.

This is, of course, a very simplified version of the professional methods of creating a cladogram. To explain what you're looking at: the systematist first gets a collection of organisms, in any order. The characters of the organisms, such as the presence of jaws, legs, etc are catalogued and put in a table like on the left. because common descent exists for these organisms we find that this list of characters can be ordered in such a way that everything above the first appearance of a character possesses that character. In this example, we can make a group of organisms based on whether they have a vertebral column. That's not interesting on it's own, but then we find an interesting phenomenon emerges: each character is found only within, and never outside a given group. Jaws are found ONLY in the vertebrate group. Legs are found ONLY in the jawed group. Amniotes are found ONLY in the legged group, and so on.
Because of this, the cladogram can be constructed as shown. There is only one correct way to draw this cladogram, because all characters are always bound within a parent group. Leading neatly to the answer to your last question, this phenomena of groups found exclusively within other groups is a specific kind of hierarchy that is exhibited only by things produced by a bifurcating pattern of descent: you could never get your books to fit a true nested hierarchy. To do so would mean for example, that you could first divide your books into hardcover and softcover. Then, you would find that red books can be found ONLY in the hardcover group, and green ones only in the softcover, with no exceptions. Further, you would find, for example, that history books are found only in the red group, while fiction is found only in the green group.
Of course, this is impossible for books. That it IS possible for organisms is very strong evidence for common descent. The reason for this is that common descent explains why we find this pattern very well. To expound: look again at the cladogram above. As a relationship tree, the y-axis is time, and where a branch becomes two is a speciation. Thus, if you could go back in time to where tuna speciated, you can draw a horizontal line across the diagram to see what species would have lived at the time. You can see that at that time, there were lancelots and lampreys, but no tigers, turtles or salamanders.
With that in mind, here is how the hypothesis of common descent explains this nested pattern. Lancelots speciated before the vertebral column evolved in the sister lineage. Lampreys inherited the vertebral column, and then speciated. Jaws evolved in the other lineage, and everything else that would eventually evolve in that lineage inherited jaws.
You can probably see where this is going. To put it another way, just in case my explanation stinks, common descent explains this pattern by hypothesising that everything with four walking legs descends from a common ancestor that also had jaws: therefore everything with 4 legs will also have jaws, but organisms with jaws could comprise a larger group in which some but not all members will have legs. This is the essence of nested hierarchies, and is why only things with some form of common descent can be placed in such a hierarchy.
The case for common descent can be made even stronger by introducing the findings of molecular biology. I will spare you all for now, lest I bore the lot of you half to death![/img]
-
otseng
- Savant
- Posts: 20518
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #4
Why would an organism then not change after millions of years? How can it be explained that some would evolve and some would basically be the same form as millions of years ago? How were they able to escape mutations for millions of years? (Horseshoe Crab is one example)Didymus wrote: All organisms in the lower part of the tree are dead, but not necessarily extinct. Remember that the phylogenetic tree is very like a family tree: everyone at the top of the tree is a modern person, while people further down are ancestors. Just because a species can be found in the lower branches of the tree doesn't mean you can't still find them alive today: all this means is that there is a branch extending from the ancient species all the way to the top.
However, looking at this tree, there is one major characteristic that have independently evolved over multiple branches - birth methods.That's not interesting on it's own, but then we find an interesting phenomenon emerges: each character is found only within, and never outside a given group.
Mammals are mostly viviparous (live-birth), however, there are exceptions that are oviparous (egg-laying), such as the Platypus.
Most reptiles are egg-laying, with some exceptions (vipers).
Most fish are egg-laying, with some exceptions (shark).
Most amphibians are egg-laying, with also some exceptions.
So, here is one major characteristic that cannot be classified within a single group. Therefore it violates the true nested criteria that you mention.
Post #5
Excellent point, my friend. Now, we've stumbled on to another interesting aspect of systematics: separating homology from analogy.
In systematics, a distinction is made between traits which we think are derived from a common ancestor (called homologous traits) and those that evolved independently (called analogous). This is a vital distinction because, as you've noted, only homologous traits are expected to fall into nested hierarchy.
Fortunately, it's usually very easy to tell if a trait is homologous. Both mammals and insects have eyes, for example, but if we look closely it's easy to tell that they are not anything like the same kind of eye. Insect eyes are like a mass of telescopic tubes, while mammal eyes are totally different, more like a camera. By analysing the anatomy of a character in detail, rather than a cursory glance, it becomes easy to tell what traits are truly related to one another.
For an obvious example, both plants and some parrots are green, but they certainly don't belong in a 'green things' group together. At first glance this may violate nested hierarchy, but in fact if we look closely, the green of plants and parrots is totally different. Plants are green due to chlorophyll, the molecule that allows them to absorb sunlight for energy, while parrots haven't got a molecule of chlorophyll in them: they are green because of an entirely different kind of pigment molecule. Because of this significant difference, greenness of plants and parrots is considered an analogous trait and should be ignored in making cladograms.
I'll come back to that in just a moment, but first I want to point out something slightly different in the case of your mammal example.
I have taken this image from the excellent Tree of Life systematics website (www.tolweb.org). I have added the dot and box you see in blue.

Nested hierarchy does not mean that our named groups such as mammals should share all characters. Instead, it means that for any character, we should be able to pinpoint a dot on the cladogram where the trait first evolved, and then everything above the dot will have the trait, and nothing else. Imagine the tree made of glass tubes with water flowing toward the 'top' of the tree (or the side, wherever the modern species are listed). you should be able to put all organisms with a given trait into one group by putting a drop of coloring into the tree at one point.
So, in the example above, we can group all mammals with live birth by putting blue dye in the tree at the point I've shown. As you can see, it's not a violation of cladistics for monotremes to lay eggs, because the viviparous group starts after the branching-off of the monotremes.
Now, where were we. As you've noticed, there are traits that seem to be out of place. Some sharks are also viviparous, yet they are supposed to be well and truly outside the mammal group. You can not put dye into your tree in any way that includes all viviparous species and no non-viviparous species.
So, the next step is to decide whether all viviparous characters are homologies, or whether the traits actually differ from one another, like parrot and plant greennesses.
To do that, we have to look at the trait in detail: just how much like mammalian viviparity is shark viviparity? When we look at this, we find something we could have predicted already: the evidence suggests that viviparous sharks evolved their very own method that is significantly different from that in mammals. First off, we find that those sharks that are considered most primitive still lay eggs. (Important note: in systematics this does not imply an advancement, like in 'primitive technology'. Instead, it is a more precise term referring to how far down the tree the group split from ancestors. In this case it means those sharks that represent species that split from other sharks soon after the split from other fish). If common descent is true, these sharks are expected to have more in common with fish, so the fact that they have eggs is pretty telling on its own.
That's far from the least of it, though: even more evidence for the idea that sharks evolved live birth independindependentlyrom the large group of sharks that emply a employductive strategy called OVOVIVIPARITY, which as the name suggests, is actually an intermediate form between egg laying and live birth. The shark has eggs alright, but they are modified, membranous ones, and they are not laid but kept in the uteris until well formed embryos can be expelled. We can even tell why the shark may have evolved this strategy: the embryos are protected during the most vulnerable part of their development, and can come into the world big, toothy, and fighting fit.
What we're looking at is an extant intermediate form, or a living species that is descended from an ancestral species that also gave rise to more advanced forms. In this case, the most recently evolved sharks, the hammerheads, have true viviparity. The evidence, of course, suggests that this gradually evolved from the ovovivovoviviparitye less recently evolved shark families, which in turn evolved from an ancestral shark species that laid their eggs in the traditional method, and is represented today by ancestors in which we can still see this primitive character.
Importantly, and very tellingly, the placental viviparity in hammerheads is significantly anatomically different from the same method in mammals, which would suggest that the trait is analogous on its own, even without all this extra evidence that gives us a good picture of how sharks invented viviparity.
So, the science of systematics led us to expect that viviparous sharks evolved the trait inindependentlyand that it would turn out to be quite different from the viviparity found in mammals. Armed with this, we find that not only is shark viviparity very different as expected, but there is a bunch of evidence giving us a good idea of how and when it evolved. Systematics is thus fully justified in doing what it does in this situation: it labels shark viviparity and mammal viviparity as two separate characters, reevaluates the cladograms, and finds that nested hierarchy prevails.
I have not had time to investigate vipers or amphibians, but I expect a similar story. Their viviparous characters will turn out to be very different from the mammal version. By the way, do you have any idea which amphibians are supposed to do this?
In systematics, a distinction is made between traits which we think are derived from a common ancestor (called homologous traits) and those that evolved independently (called analogous). This is a vital distinction because, as you've noted, only homologous traits are expected to fall into nested hierarchy.
Fortunately, it's usually very easy to tell if a trait is homologous. Both mammals and insects have eyes, for example, but if we look closely it's easy to tell that they are not anything like the same kind of eye. Insect eyes are like a mass of telescopic tubes, while mammal eyes are totally different, more like a camera. By analysing the anatomy of a character in detail, rather than a cursory glance, it becomes easy to tell what traits are truly related to one another.
For an obvious example, both plants and some parrots are green, but they certainly don't belong in a 'green things' group together. At first glance this may violate nested hierarchy, but in fact if we look closely, the green of plants and parrots is totally different. Plants are green due to chlorophyll, the molecule that allows them to absorb sunlight for energy, while parrots haven't got a molecule of chlorophyll in them: they are green because of an entirely different kind of pigment molecule. Because of this significant difference, greenness of plants and parrots is considered an analogous trait and should be ignored in making cladograms.
I'll come back to that in just a moment, but first I want to point out something slightly different in the case of your mammal example.
I have taken this image from the excellent Tree of Life systematics website (www.tolweb.org). I have added the dot and box you see in blue.
Nested hierarchy does not mean that our named groups such as mammals should share all characters. Instead, it means that for any character, we should be able to pinpoint a dot on the cladogram where the trait first evolved, and then everything above the dot will have the trait, and nothing else. Imagine the tree made of glass tubes with water flowing toward the 'top' of the tree (or the side, wherever the modern species are listed). you should be able to put all organisms with a given trait into one group by putting a drop of coloring into the tree at one point.
So, in the example above, we can group all mammals with live birth by putting blue dye in the tree at the point I've shown. As you can see, it's not a violation of cladistics for monotremes to lay eggs, because the viviparous group starts after the branching-off of the monotremes.
Now, where were we. As you've noticed, there are traits that seem to be out of place. Some sharks are also viviparous, yet they are supposed to be well and truly outside the mammal group. You can not put dye into your tree in any way that includes all viviparous species and no non-viviparous species.
So, the next step is to decide whether all viviparous characters are homologies, or whether the traits actually differ from one another, like parrot and plant greennesses.
To do that, we have to look at the trait in detail: just how much like mammalian viviparity is shark viviparity? When we look at this, we find something we could have predicted already: the evidence suggests that viviparous sharks evolved their very own method that is significantly different from that in mammals. First off, we find that those sharks that are considered most primitive still lay eggs. (Important note: in systematics this does not imply an advancement, like in 'primitive technology'. Instead, it is a more precise term referring to how far down the tree the group split from ancestors. In this case it means those sharks that represent species that split from other sharks soon after the split from other fish). If common descent is true, these sharks are expected to have more in common with fish, so the fact that they have eggs is pretty telling on its own.
That's far from the least of it, though: even more evidence for the idea that sharks evolved live birth independindependentlyrom the large group of sharks that emply a employductive strategy called OVOVIVIPARITY, which as the name suggests, is actually an intermediate form between egg laying and live birth. The shark has eggs alright, but they are modified, membranous ones, and they are not laid but kept in the uteris until well formed embryos can be expelled. We can even tell why the shark may have evolved this strategy: the embryos are protected during the most vulnerable part of their development, and can come into the world big, toothy, and fighting fit.
What we're looking at is an extant intermediate form, or a living species that is descended from an ancestral species that also gave rise to more advanced forms. In this case, the most recently evolved sharks, the hammerheads, have true viviparity. The evidence, of course, suggests that this gradually evolved from the ovovivovoviviparitye less recently evolved shark families, which in turn evolved from an ancestral shark species that laid their eggs in the traditional method, and is represented today by ancestors in which we can still see this primitive character.
Importantly, and very tellingly, the placental viviparity in hammerheads is significantly anatomically different from the same method in mammals, which would suggest that the trait is analogous on its own, even without all this extra evidence that gives us a good picture of how sharks invented viviparity.
So, the science of systematics led us to expect that viviparous sharks evolved the trait inindependentlyand that it would turn out to be quite different from the viviparity found in mammals. Armed with this, we find that not only is shark viviparity very different as expected, but there is a bunch of evidence giving us a good idea of how and when it evolved. Systematics is thus fully justified in doing what it does in this situation: it labels shark viviparity and mammal viviparity as two separate characters, reevaluates the cladograms, and finds that nested hierarchy prevails.
I have not had time to investigate vipers or amphibians, but I expect a similar story. Their viviparous characters will turn out to be very different from the mammal version. By the way, do you have any idea which amphibians are supposed to do this?
-
Oolon Colluphid
- Student
- Posts: 13
- Joined: Tue Jun 29, 2004 6:41 am
- Location: Hampshire, England
Post #6
And cyanobacteria are an even better one. If I say to you the oft-heard creationist claim, ‘if we evolved from monkeys, why are there still monkeys?’, do you start to see the answer?otseng wrote:Why would an organism then not change after millions of years? How can it be explained that some would evolve and some would basically be the same form as millions of years ago? How were they able to escape mutations for millions of years? (Horseshoe Crab is one example)
It is because, once you have a phenotype that is well-suited to a particular niche, there is little selection pressure to deviate from that phenotype. In fact, moving away from that form is likely to be punished by natural selection... unless doing so allows success in a different niche. Morphology tracks the requirements of the niche. So if the niche is stable over a long period, so will the phenotype be.
The monkey shape is an effective shape to be in arboreal niches... but not so good in a brain-power-and-tool-using niche. The horseshoe crab and coelacanth shapes are good ‘designs’ within their own niches. And bacteria do a good job of being bacteria. In effect, there are countless ‘living fossils’ -- beetle-shaped ones, fish-shaped ones, ant-shaped ones, nautilus-shaped ones, monkey-shaped ones, lizard-shaped ones, and so on. But the particular species and genera we now have are not the same as the fossil ones.
Note that it is not the case that ‘living fossils’ have not changed at all, let alone in their genetics (if only we could compare the ancient versions!). They haven't escaped mutations, just most of the mutations that would change them morphologically, because that would change how well they molded to their niches. The modern, classic ‘living fossil’, the coelacanth for instance, is in the faily Latimeriidae, whereas fossil ones are family Coelacanthidae, and differ from the living genus in a number of ways, including size and various internal structures. Note though that their deep ocean habitat has been stable for a very long time.
Note that all of those are vertebrates, and all seem to have the basic idea of retaining the egg in the body rather than laying it. I don’t know of any viviparous insects! But as Didymus points out, the difference is in the detail. Some viviparous shark species, for instance, even have a form of placenta. However, each embryo has its own separate placenta, whereas the mammalian one is (please correct me if wrong, this is off the top of my head) a single one for all the foetuses.However, looking at this tree, there is one major characteristic that have independently evolved over multiple branches - birth methods.
Monotremes -- the platypus and echidnas -- lay eggs... but they also have shared derived characteristics that link them with mammals -- hair, lactation, etc. The problem here is, in cladistic terms, with deciding the polarity of the characteristics. And we do this by comparing them to an out-group (here, pick any other vertebrate group... they generally lay eggs). Therefore milk and fur link the monotremes with the marsupials and placentals, to the exclusion of other vertebrates.
Sure, convergent evolution can confound the pheneticis. Which is why most folks these days go with cladistic analyses of character sets!
Cheers, Oolon
-
otseng
- Savant
- Posts: 20518
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #7
From my understanding of evolution, natural selection has nothing to do with genetic mutations. They do work hand in hand to produce evolution, but natural selection does not influence whether genes are mutated or not.Oolon Colluphid wrote: It is because, once you have a phenotype that is well-suited to a particular niche, there is little selection pressure to deviate from that phenotype. In fact, moving away from that form is likely to be punished by natural selection... unless doing so allows success in a different niche. Morphology tracks the requirements of the niche. So if the niche is stable over a long period, so will the phenotype be.
Therefore, natural selection cannot explain why organisms have not mutated over millions of years.
Last edited by otseng on Thu Jul 08, 2004 10:10 pm, edited 1 time in total.
-
otseng
- Savant
- Posts: 20518
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #8
Is there an objective test available to determine whether a trait is homologous or analogous?Didymus wrote: In systematics, a distinction is made between traits which we think are derived from a common ancestor (called homologous traits) and those that evolved independently (called analogous). This is a vital distinction because, as you've noted, only homologous traits are expected to fall into nested hierarchy.
Also, the tree graph I presented above was the most comprehensive one I could fine. Do you (or anyone else) have a more comprehensive diagram of a phylogenetic tree? Perhaps it would be more beneficial to the debate if we can work off the best possible source.
I have no idea of the specific names of the amphibians that are viviparous. However, here is my source of the animals I cited.I have not had time to investigate vipers or amphibians, but I expect a similar story. Their viviparous characters will turn out to be very different from the mammal version. By the way, do you have any idea which amphibians are supposed to do this?
Post #9
There is not A single test, because the question is far too complicated for codification. Rather, the separation of homology from analogy is one of the most often dealt with subjects in the field of systematics. In other words, many scientists devote their entire professional careers to just this question alone, and objectively establishing the answers is exactly what they strive for. In many cases the answer is easy, such as the investigation of sharks that I demonstrated above. In other cases, particularly with very minute characters in very closely related species, the problem is much more difficult to solve and there is controversy. Of course, controversy does not mean that the question is subjective, just that it hasn't been decided to everyones satisfaction.otseng wrote:Is there an objective test available to determine whether a trait is homologous or analogous?
Furthermore, the areas of controversy are usually in the highest parts of the tree: trying to establish the precise relationships of three very similar species of moth, for example, would be difficult to do because this method relies on the evolution of new characters after a split. If the split occurred very recently then there will be very few characters that have evolved in the meantime. Establishing the relationships of major groups is usually much easier, and there is little controversy.
One of my favourite websites is Tolweb, from which I obtained my marsupial relationships diagram. The site is a series of small diagrams all linked together like a book of roadmaps, so that the tree can go into much greater depth than any one diagram could be large enough for.Also, the tree graph I presented above was the most comprehensive one I could fine. Do you (or anyone else) have a more comprehensive diagram of a phylogenetic tree? Perhaps it would be more beneficial to the debate if we can work off the best possible source.
www.tolweb.org
I'm currently looking as some extremely interesting work on vipers. More soon.
Post #10
This is great stuff. I've just had a whale of a time looking into the evolution of snake babies, which may be a bit perverse, but what the heck.
A great deal of the life work of one D.G. Blackburn has gone into the question of independently evolved viviparity.
From Blackburn, D.G. (1995). Saltationist and punctuated equilibrium models for the evolution of viviparity and placentation. Journal of Theoretical Biology 174: 199-216:
Lest it be thought that this "evolved independently" idea is just an ad-hoc attempt to explain away what ought to be problematic incongruences, I offer this particularly interesting paper:
Shine, R. 2002. Reconstructing an adaptationist scenario: what selective forces favor the evolution of viviparity in montane reptiles? American Naturalist 160:582-593.
This is an extremely interesting paper in it's own right, dealing particularly with the question of adaptationist scenarios and their testability. However, like most scientific papers it's of interest only to devotees of the topics it deals with. I love it, for example, and I'm sure my friend Oolon here will also enjoy it. However, I won't burden anyone with the responsibility of reading the whole thing just to make my point in this discussion.
Instead, I'll draw out the important information from the paper:
Veeery fun stuff. Thankyou for pointing me in this direction.
A great deal of the life work of one D.G. Blackburn has gone into the question of independently evolved viviparity.
From Blackburn, D.G. (1995). Saltationist and punctuated equilibrium models for the evolution of viviparity and placentation. Journal of Theoretical Biology 174: 199-216:
Because of this dichotomy, says blackburn, the transition from oviparity to some form of live birth proceeds as a gouldian punctuation: the species will retain oviparity until the scales are finally tipped and the costs outweigh the benefits. When that happens, the transition happens very quickly (in evolutionary terms). Blackburn identifies over 100 reptile species in which viviparity is thought to have evolved independently.Evolution in squamates seems to have produced a dichotomy between two evolutionarily stable patterns: 1) retention of weakly-shelled or shell-free eggs to term (viviparity), with development of fully functional placentae; and 2) deposition of shelled eggs at or near the limb bud stage of development (typical oviparity). Conflicting functional demands placed on eggshell morphology may constrain establishment of prolonged, oviparous egg-retention as a viable, historically stable pattern. Alternatively, the costs of prolonged egg-retention associated with decreased female mobility or decreased fecundity may exceed the benefits in oviparous forms.
Lest it be thought that this "evolved independently" idea is just an ad-hoc attempt to explain away what ought to be problematic incongruences, I offer this particularly interesting paper:
Shine, R. 2002. Reconstructing an adaptationist scenario: what selective forces favor the evolution of viviparity in montane reptiles? American Naturalist 160:582-593.
This is an extremely interesting paper in it's own right, dealing particularly with the question of adaptationist scenarios and their testability. However, like most scientific papers it's of interest only to devotees of the topics it deals with. I love it, for example, and I'm sure my friend Oolon here will also enjoy it. However, I won't burden anyone with the responsibility of reading the whole thing just to make my point in this discussion.
Instead, I'll draw out the important information from the paper:
In other words, Professor Shine has not only constructed a robust adaptationist hypothesis for the observed frequency of individual reptilian 'inventions' of viviparity, but was able to objectively test this hypothesis. He found that there is a certain temperature boundary, associated with an altitude in this study, at which the advantages of retaining eggs suddenly take a big jump. This is strong evidence in support of Blackburn, who's punctuated equilibrium model of reptilian viviparity would predict exactly this sort of jump in fitness requirements at a certain boundary.This study provides the most direct information yet available on selective forces operating on the uterine retention of eggs (and, thus, on the evolution of viviparity) ...
My data strongly support all of the following predictions from the “cold-climate” hypothesis for the evolution of reptilian viviparity. First, the upper elevational limit for egg layers in the field corresponded to the elevation above which there was a marked decrease in hatching success and hatchling viability. Second, this viability decrease at higher elevations comprised both a reduction in hatching success and a decrease in mean values for several hatchling traits (body size, growth rate, running speed) likely to affect organismal fitness (Andrews 2000). Third, brief retention at higher temperatures (mimicking those experienced by eggs in utero) enhanced both hatching success and offspring viability. Fourth, the viability increment due to “uterine” retention was greater for eggs incubated for the rest of development at cooler rather than warmer temperatures in the laboratory and at higher rather than lower elevations in the field. And, fifth, progressive increases in the duration of uterine retention generated increasing benefits to offspring viability. Thus, in the absence of constraints or opposing disadvantages, we would expect selection to favor a progressive increase in the duration of uterine retention of developing eggs at elevations above the usual upper limit for oviparous reproduction.
Veeery fun stuff. Thankyou for pointing me in this direction.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}