

As per the request in this thread, I am starting this thread to debate the question of common descent. I'm perfectly happy to kick off the discussion with a presentation of a tiny slice of the evidence biologists consider established common descent.
Common descent is a historical claim about the evolutionary past of organisms. It refers to the idea that all organisms are related to one another like a gigantic extended family. For example: five million years ago, chimpanzees and humans did not exist separately, but were represented by a single species. Approximately 4.5 million years ago that population underwent speciation and each new species (at that point very similar to one another), gradually diverged. According to all of mainstream biology, this principle holds true for all life, so that every species shares ancestry with every other at some point in the distant past.
To make this claim, biologists have a large amount of evidence that they think establishes common descent as a very robust scientific claim. The famous talkorigins '20+ evidences for macroevolution' article is actually a list of evidence for common descent, and doesn't actually have much to do with macroevolution as modern biology uses the term.
Rather than use that article here, I will instead go strait to the most authoritative sources I have access to, and give a quick summary of the overview of the evidence from Douglas J. Futuymas Evolutionary Biology 3rd edition, the most respected general textbook in the field.
Futuyma lists eight categories of evidence, each of which would require a fair amount of elaboration. I'll give the full list before expanding on one or two.
1. The hierarchical organisation of life.
2. Homology
3. Embryological similarities
4. Vestigial characters
5. Convergence
6. Suboptimal design
7. Geographical distributions
and 8. Intermediate forms.
I'll expand a little on 1 and 8 before throwing open the floor.
Life on earth exists in a nested hierarchy. This means that if you divide organisms into groups according to their features, you can then divide each of those groups into more groups without overlapping. So, if one divides the tetrapods from the arthropods, one can then pick out smaller groups; say, the mammals out of the tetrapods, and the insects from the arthropods, and those groups will not overlap in any new features that are not present in their parent groups. Meanwhile, if you try to group cars first by engine size, and then by colour, you'll find that there is a lot of overlap of features in groups that aren't present in those groups parent categories. So both red cars and blue cars can have both V6 and V12 engines, but triple segmentation with six legs is found only in arthropod groups (namely insects), while milk is found only in tetrapod groups (namely mammals).
This is evidence for common descent, because this pattern of nested hierarchy is produced by a historic pattern of descent and divergence, and by hardly any other types of processes. For example, tracing the surnames in a family tree yields a nested hierarchy: if one branch of the plantaginet family changes its name to plantaginet-smith, then you'll find plantaginet smith only in the larger plantaginet extended family. Only objects that have been produced by descent and branching fall into true nested hierarchies.
This fact about living things allows us to do all sorts of things in the exciting world of molecular biology. Nested hierarchies are what enable us to produce phylogenetic trees of life, which are essentially hypotheses about a particular pattern of descent. Using the same methods we now use to establish paternity from blood samples in court, we establish the relationships of species. The most striking thing is that we can yield the same phylogenetic tree by analysing just about any genetic feature: be it a gene, or mitochondrial DNA, or a particular protein sequence like hemoglobin, or most tellingly, endogenous retroviral insertions (I can expand on this last one if it's unfamiliar), the same tree keeps appearing again and again. This would be inexplicable if all species had separate origins, but is exactly what common descent predicts.
Now, to intermediate forms. Here, I will list the fossilised forms of three major evolutionary lineages, from three of my favourite books. I can't supply the diagrams, obviously, but I will give page numbers for anyone who wants to check up on me.
First, it should be established that according to the scenario of independent creation of organisms, intermediate sequences should not exist at all. Given that, the following three gradual evolutionary sequences establish the evolutionary history of these organisms: lizards to mammals, four-legged artiodoctyls to whales, and hyracotherium to modern horses.
From Ridleys Evolution text: The following fossils form a transitional lineage: Reptilian pelycosaurs to Ophlacodontids (which also gave rise to the famous Dimetrodon ), to the broadly varying Therapsids, from amongst whos number came the Cynodots, representing whom there are many individual specimens, such as(in approximate evolutionary order):
Procynosuchus
Thrinaxodon (here's another thrinaxodon)
Diademodon
Probainognathus
These specimens become increasingly less reptilian and begin to accumulate mammilian features over time. The latest cynodonts in this series blend neatly into the early mammals.
I'm running short on time, so I'll finish whales and horses quickly.
Refer to At the Waters Edge by Carl Zimmer for more information on this graduation from four legged pakicetus to modern whales:
Pakicetus, Ambulocetus, Dalanistes, Rodhocetus, Takracetus, Gaviocetus, Basilisaurus, Dorudon, Mysticetes (modern baleen whales). Googling these names might yield more information as well.
Refer to Strickberger and Monroes' Evolution textbook for more information on the transition from the small 55 million year old Hyracotherium, through the Oligocene Miohippus and Miocene Merychippus to modern Equus. This is a famously well documented transition, so it should be easy to find online resources.
But right now, I've made myself late. I suppose the topic of this debate should be: 'Do these observations, which are commonly advanced by mainstream biology, provide evidence for common descent?'
Good night and enjoy the debate.
The reality or otherwise of Common Descent
Moderator: Moderators
-
otseng

- Savant
- Posts: 20517
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #41
If common descent is true (and the phylogenetic tree is true), it would be expected that morphology and molecular systematics agree with each other. Yet, the article shows that this is not the case. Sure, the purpose of the article was not to disprove common descent. But, it does bring into question how the phylogenetic tree is to be constructed.fried beef sandwich wrote:My response: The article is talking about the relative merits of two techniques and systems of evolutionary organization. Common descent itself is not in debate.otseng wrote:Another question about the phylogenetic tree, how do you respond to allegations that morphology and molecular systematics produce differing results? Bones, molecules, ... or both?
The issue is the explanation of new morphological features. Microevolution can explain antibiotic resistance. And there is no dispute about microevolution.fried beef sandwich wrote: Bacteria can gain antibiotic resistance. Sure, it's still a single-celled organism, but I'm just putting it there to show you that gaining features can be done.
However, it's alterability does not indicate whether the model is valid or not.
The ability of a model to be open to the possibility of change in light of new evidence is a strength, not a weakness.
-
otseng
- Savant
- Posts: 20517
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #42
I will bring some evidence up when I start talking about intelligent design. (It might be awhile though since there are several active threads in C vs E.)ST88 wrote: Of all the discoveries that have been made, nothing points away from evolution or common descent.
Also, I think we are starting to stray a bit from the main point of this topic. The main issue is, "what are the evidence for and against common descent?" Whether the model is alterable or not does not provide any evidence for it's support. So I ask, what is the evidence for the validity of common descent?
-
fried beef sandwich

- Student
- Posts: 49
- Joined: Sat Jun 26, 2004 1:59 am
- Location: Southern California
Post #43
Your logic is right, but your argument is all wrong. This is your logic:otseng wrote:If common descent is true (and the phylogenetic tree is true), it would be expected that morphology and molecular systematics agree with each other. Yet, the article shows that this is not the case. Sure, the purpose of the article was not to disprove common descent. But, it does bring into question how the phylogenetic tree is to be constructed.
1. [Common Descent (true) && Tree (true)] --> (Morph && Mol)
2. ~(Morph && Mol)
3. ~[Common Descent (true) && Tree (true)]
And when you critique the logical propositions as such, the whole thing works as an example of an indirect proof.
The problem is this: your first premise is wrong.
whether or not molecular systematics or morphological systematics agree with each other has no bearing on whether or not all organisms have descended from a common ancestor. If they disagree, all it means is that the current phylogenetic tree is written incorrectly in some way.
Example: the horseshoe crab is more closely related to a spider than a crab. Does this mean that it did not evolve from an earlier life form? No. Does it mean it does not have a common ancestor with some other life form? No. Does this mean that evolution is now a theory in crisis? No. All it means is that it's been classified it incorrectly. A mango sitting in the orange section is still a tropical fruit.
So, your argument should have read:
(1) Tree (true) --> (Morph && Mol)
(2) ~(Morph && Mol)
(3) The phylogenetic tree is flawed in some way.
Like I said, the issue of whether or not organisms descended from earlier, more primitive ancestors has nothing to do with what the phylogenetic tree should look like.
What do you mean by "new morphological features" anyway? At what scale does a mutation qualify as a "new morphological feature"? Please explain. It's very hard to debate constructively when you're trying to cast things around in vague, undefinable terms.The issue is the explanation of new morphological features.
On the bacterial scale of things, a mutation that causes your cell wall to be impregnated by lipoproteins and ligands, hence rendering it well-nigh impenetrable by antibiotic compounds definitely qualifies as a "new morphological feature." For a human, that's the equivalent of suddenly growing a one-inch thick layer of rock-hard skin.
For a bacteria that mutates and is now able to manufacture a new enzyme that allows it to digest a previously unexploitable food source, that's like getting an opposable thumb or another pair of hands.
Or, how about a bacteria that mutates to form a flagella with a different microtubule configurations than its previous generation? On the human scale, that's like growing T-rex arms instead of your normal hands.
Or maybe you want to see a transition between unicellular and multicellular life forms? That's why I brought up the Volvox colonies. Have you checked those little buggers out yet? They're wee little beasties that live in spherical colonies, reproducing via daughter colonies.
Microevolution can explain antibiotic resistance. And there is no dispute about microevolution.
Here's what I don't understand: how can you believe in "microevolution" but not "macroevolution"? That's like saying you don't believe that the answers you get in calculus are true because, although you 'believe in' estimating the area under a certain section of the graph, you don't believe that you can use the same technique, refined many times, to find the area under the entire graph.
Macroevolution is the calculus of microevolution. It's what you get when you add up all the mutations in an organism that is microevolving over millions of years. Everything macroevolves. Populations that are microevolving are macroevolving as well.
-
otseng
- Savant
- Posts: 20517
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #44
From the very first post in this thread, it is brought up that one of the evidence that common descent is true is "the hierarchical organisation of life". One of the exhibits that was brought forth was the phylogenetic tree to demonstrate this organization. So, my argument is showing that there is no consistent, objective method on how this phylogenetic tree is to be constructed. Since the validity of the phylogenetic tree is in dispute, then it is not good evidence to support common descent.fried beef sandwich wrote: whether or not molecular systematics or morphological systematics agree with each other has no bearing on whether or not all organisms have descended from a common ancestor. If they disagree, all it means is that the current phylogenetic tree is written incorrectly in some way.
So, your argument should have read:
(1) Tree (true) --> (Morph && Mol)
(2) ~(Morph && Mol)
(3) The phylogenetic tree is flawed in some way.
For example, I would clasify the cilium as a new morphological feature. It is a new physical feature that adds new functionality (propulsion) to a cell.What do you mean by "new morphological features" anyway? At what scale does a mutation qualify as a "new morphological feature"? Please explain. It's very hard to debate constructively when you're trying to cast things around in vague, undefinable terms.The issue is the explanation of new morphological features.
I have visited the site you presented. What exactly is the Volvox a transition between?Or maybe you want to see a transition between unicellular and multicellular life forms? That's why I brought up the Volvox colonies. Have you checked those little buggers out yet? They're wee little beasties that live in spherical colonies, reproducing via daughter colonies.
We could probably argue about macro vs micro with no end in sight. That's why this thread was spawned off of the macro vs micro debate.Here's what I don't understand: how can you believe in "microevolution" but not "macroevolution"? That's like saying you don't believe that the answers you get in calculus are true because, although you 'believe in' estimating the area under a certain section of the graph, you don't believe that you can use the same technique, refined many times, to find the area under the entire graph.
I believe the analogy is more like estimating y at x=100 when all we know is that x=0, y=0 and x=1, y=2.
It's a simple concept to say, but difficult to demonstrate.Macroevolution is the calculus of microevolution. It's what you get when you add up all the mutations in an organism that is microevolving over millions of years. Everything macroevolves. Populations that are microevolving are macroevolving as well.
I suggest that perhaps the best line to progress in this dialogue is what Didymus laid out in the very first post. He presented 8 arguments for the support of common descent (none of which uses micro/macroevolution). This gives a good starting point on the evidence for and against common descent.
I believe we have covered the first two:
1. The hierarchical organisation of life.
2. Homology
If someone would like to add more to these two, please do.
However, I would like to hear evidence from the third:
3. Embryological similarities

Post #45
Just to put this into plain English, the phylogenetic tree based on morphology predates and operates outside of genetic testing. The fact that the two systems of classifications are similar but not exact only points to their opposition. The real controversy is that morphologists, in this sense, are clinging to their research in the face of genetic evidence because they don't want their lives to have been wasted. The fact that they are now questioning genetic testing because of the way the evidence is being interpreted I think belies their true motives.otseng wrote:If common descent is true (and the phylogenetic tree is true), it would be expected that morphology and molecular systematics agree with each other. Yet, the article shows that this is not the case. Sure, the purpose of the article was not to disprove common descent. But, it does bring into question how the phylogenetic tree is to be constructed.
However, neither side is in disagreement about the basic theories behind evolution, whether convergent or progressive or both. The fact that all primates had a common ancestor is not in dispute, only whether or not existing species, such as Chimpanzees, pre-figured humans or were merely alongside them and the actual human line is lost to history (so far).
I think I see where you're going with this, and I have to disagree. The morphological similarity angle could very well be convergent evolution in practice, but it's highly unlikely. The genetic similarity makes it harder to claim convergent evolution because there would be many more genetic differences, if not morphological differences, than have been currently found. We can see semi-convergent evolution in sharks and dolphins, for example. But no one will claim that the set of genes that produces the shark dorsal fin are the same as the set that produces the dolphin dorsal fin.
Antibiotic resistance has to do with evolution in only one way, and it has nothing to do with "gaining" resistance, it has to do with natural selection. Genetic variability (not mutability) within bacteria colonies in any given location virtually assures that a very very small percentage of them will be resistant to a given antibiotic. This is from the get-go, not over time. The resistant bacteria will not respond to one antibiotic, so it becomes necessary to use successive treatments.otseng wrote:The issue is the explanation of new morphological features. Microevolution can explain antibiotic resistance. And there is no dispute about microevolution.fried beef sandwich wrote: Bacteria can gain antibiotic resistance. Sure, it's still a single-celled organism, but I'm just putting it there to show you that gaining features can be done.
However, the problem now is that these single-antibiotic resistant bacteria have become so prevalent that they are now the pool from which other bacteria form, and the inherent genetic variability (not mutability) in the species says that a very very small percentage of those that do not respond to penicillin will now also not respond to erythromycin. And so on.
I agree with you about validity. But viability is a different thing. I concede that your standards of 99.999% genetic similarity are far too high to constitute requisite proof, but doubt is sometimes more powerful than evidence. And the hyperbolic approach of the evidence towards that number must necessarily increase doubt inversely. My point was that evidence supporting common descent is only growing and will never decrease because of its alterability. This does not necessarily decrease the evidence for any other competing theory, but as one becomes stronger, the other necessarily weakens.otseng wrote:However, it's alterability does not indicate whether the model is valid or not.
The theories are not bound to the interpretations of the evidence by researchers. We can all look at the facts and come up with different algorithms for their existence. Within the scope of common descent, some theories work and some don't. But just like natural selection, those that best fit the evidence tend to gain support and those that don't tend to die off. Viability is not affected in this case because the over-arching idea of common descent is not in dispute.
-
otseng
- Savant
- Posts: 20517
- Joined: Thu Jan 15, 2004 1:16 pm
- Location: Atlanta, GA
- Has thanked: 197 times
- Been thanked: 337 times
- Contact:
Post #46
ST88 wrote:The fact that all primates had a common ancestor is not in dispute
Perhaps not in dispute by evolutionists. But it is certainly in dispute by creationists.
Antibiotic resistance has to do with evolution in only one way, and it has nothing to do with "gaining" resistance, it has to do with natural selection. Genetic variability (not mutability) within bacteria colonies in any given location virtually assures that a very very small percentage of them will be resistant to a given antibiotic. This is from the get-go, not over time. The resistant bacteria will not respond to one antibiotic, so it becomes necessary to use successive treatments.
I stand corrected then. This leads one to wonder - how exactly does one determine if genetic mutations comes into play and that it is not simply just a result of natural selection?
My point was that evidence supporting common descent is only growing and will never decrease because of its alterability.
That assumes that evidence against common descent does not grow.
Anyways, I think arguing about the nature of the model has little value. What is important are the evidences themselves.
So, to open it up, if anyone can give evidence from any of the points in the initial post, feel free to present them. To reiterate the points, they are:
1. The hierarchical organization of life
2. Homology
3. Embryological similarities
4. Vestigial characters
5. Convergence
6. Suboptimal design
7. Geographical distributions
8. Intermediate forms
Post #47
1. The hierarchical organization of lifeotseng wrote:So, to open it up, if anyone can give evidence from any of the points in the initial post, feel free to present them. To reiterate the points, they are:
Common descent says that simpler organisms developed first. It says that their genetic codes would be simpler and that many characters in simpler organisms would be passed on to more complex ones.
All plants and animals are made up of what are essentially eukaryotic cells, sharing many characteristics of single-celled eukaryotic organisms, including the complex sexual reprduction process. This, despite the fact that there are other types of single-celled organisms, prokaryotes, which multiply through simple cell division.
Prokaryotes have vastly smaller amounts of DNA within them and are therefore theorized to have evolved first. Despite their differences, they still rely on structures like DNA, ribosomes, and operate with ATP as energy, either absorbing it as free ATP or producing it themselves. We assume that obligate aerobes evolved into eukaryotic cells because they require oxygen. But there are prokaryotes that thrive in different environments from this.
It is important that Prokaryotes came first because the presence of one type (Cyanobacteria) increased the amount of O2 in the atmosphere by converting CO2 and H20 in seawater to CHO sugars and 02, setting the stage for obligate aerobes. These two types of organisms could not have come about at the same time because the cyanobacteria would not have been able to create enough O2 in the atmosphere for all obligate aerobes to survive right away. There would have to have been an enormous, world-wide O2 generating phenomenon for many many years as the oceans became saturated with it and it seeped into the atmosphere. The sheer amount of Cyanobacteria would have precluded many other forms of life happening at this same time. Eventually, when plants became land-bound, this would no longer be a problem as the world would no longer need the vast ocean-based 02 factory exclusively.
But mostly the heirarchy is intended to mirror the course of evolution. This will always be in dispute short of a complete and definitive analysis of the genome of all species.
2. Homology
"A similarity between species that is not functionally necessary."
Whales, bats, and cows are pentadactyls, they have five bony finger structures, yet none actually requires the use of all five.
The stingers of some wasps are actually reproductive organs, ovipositors, which explains why only females sting. Not all wasps have this feature. Most species of Cuckoo wasps and Ichneumonid wasps have the ovipositor structure, but lack the stinger adaptation.
Blind cave fish and blind cave lizards have openings in their skulls for eyes and/or the skin bulbs required to hold eyes despite the fact that they lack eyes.
The Common Dandelion reproduces asexually -- they do not pollinate themselves nor one another -- they use parthenogenesis in order to form viable seeds, yet they produce pollen and have other flower sexual structures (including the flower itself).
3. Embryological similarities
Bird embryos are homologous pentadactyls in early stages, but then lose those extra finger structures in later stages.
Unfortunately, much "evidence" in this area was fabricated in the early days of embryology by people like Haeckel, who set embryology back decades by his suspect drawings.
But there can be no dispute that embryos from fish, birds, mammals, and primates are eerily similar at early stages in their development. Whether you believe that human embryos have gill slits or not, the basic shape and formation of the embryos are the same, including notocords, pre-kidneys, and skin type.
Post #48
4. Vestigial characters
I don't believe that vestigial organs or morphologies point to common descent in any practical way. While showing that homologous structures in higher animals are useless and lower animals are useful might appear to support common descent, it equally supports the degeneration of an intelligent design of the higher animal. Common Descent nor Evolution, as theories, do not really need vestigial characters as supporting proofs. They only serve to provide accomodating theories for their existence. Further, as we learn more and more about how anatomy works, characters previously thought as vestigial have and may have become to be known as useful within the anatomy.
Similarly, vestigial genetics (the study of "junk DNA") do not support any theory any more than any other theory. The human genome has about 30,000-40,000 genes, yet there are more than 300,000 different proteins produced from this material. The complexity of this is necessarily staggering, and we have yet to truly understand how this works. But one thing is sure, the previous theory of "junk DNA" is probably false.
5. Convergence
"Organisms that are not closely related independently acquire some characteristic or characteristics in common."
Evolution says that species will adapt to their environment. Common descent says that different species will adapt in similar ways to similar environments.
The best example of this are Marsupials. There exist, or have existed, marsupial equivalents to mammals, including squirrels, badgers, mice, wolves, sloths, herding animals, bears, and other common forms according to the ecology of Australia, which has ecological analogues to other parts of the world.
We can also see this in Batesian mimicry, the classic example of which is the viceroy butterfly, which has adapted to look like the monarch butterfly to deter predators. The monarch tastes bad, so predators will avoid it based on its distinctive wing pattern, which the viceroy has successfully mimicked.
There are many other examples of this.
Personally, I find it interesting that different kinds of trees are related more closely within their "plant family" than to other trees. Apple trees, for example, are in the same family as rose bushes and strawberry vines. Yet other trees, like redbuds and orchid trees, are in the same family as lupine bushes and legume vines like beans and peas. This despite the fact that the trees more resemble each other and operate more similarly than they do to their other family members.
6. Suboptimal design
There are, in some animals, what are called "Fixed Action Patterns" or FAPs, which are kind of like hard-wired software programs in their brains. One famous example of this is a cardinal that fed fish a typical cardinal chick diet over the course of several weeks. The cardinal's nest was destroyed and its chicks killed, but the cardinal, having the instinct for feeding, took care of the fish instead. Not optimal behavior to save the species.
A FAP found in human infants is the grasping reflex. Any mother with long hair will tell you what researchers found in a controlled environment: in the presence of hair, the hands of human babies grasp more strongly than with other materials. This FAP is also seen in primates and its purpose is for the young to hold on tightly to the hair of the mother as she moves quickly. This could be a vestigial behavior instinct, but I placed it here for continuity. There is no reason for a human baby to grasp hair more strongly than other textures.
You could also argue that the flounder's life cycle makes no sense. Aside from the eye moving from one side of the head to the other throughout the juvenile form, which is explainable, why doesn't the mouth alter to be flat as well? The mouths of adult flounder are arranged sideways as if the fish were still upright. Yet this does not offer an optimal form of feeding or other mouth behaviors as horizontal mouths do in other flat animals.
7. Geographical distributions
Australia's marsupials are a good example. It would be one thing if marsupials existed only in Australia, but the opossum exists in other parts of the world as well, pointing to the evolution of marsupials as a separate group, based on their geographic isolation after opossums appeared.
Similarly, monkeys are divided into "old world" -- Africa and Asia -- and "new world" -- the Americas. Both old and new world monkeys have opposable thumbs on their feet, but only old world monkeys have truly opposable thumbs on their hands. Nostril placement on the head, number of teeth, and prehensile tails are also differences between the two populations. There are also no "ground dwelling" new world monkeys.
8. Intermediate forms
Intermediate forms do not seem to be good examples of common descent because between any two intermediate form fossils is a perceived gap of an additional intermediate form. It therefore can be argued that all intermediate forms could have arisen at the same time in the same creation event for a purpose only known to the creator.
I don't believe that vestigial organs or morphologies point to common descent in any practical way. While showing that homologous structures in higher animals are useless and lower animals are useful might appear to support common descent, it equally supports the degeneration of an intelligent design of the higher animal. Common Descent nor Evolution, as theories, do not really need vestigial characters as supporting proofs. They only serve to provide accomodating theories for their existence. Further, as we learn more and more about how anatomy works, characters previously thought as vestigial have and may have become to be known as useful within the anatomy.
Similarly, vestigial genetics (the study of "junk DNA") do not support any theory any more than any other theory. The human genome has about 30,000-40,000 genes, yet there are more than 300,000 different proteins produced from this material. The complexity of this is necessarily staggering, and we have yet to truly understand how this works. But one thing is sure, the previous theory of "junk DNA" is probably false.
5. Convergence
"Organisms that are not closely related independently acquire some characteristic or characteristics in common."
Evolution says that species will adapt to their environment. Common descent says that different species will adapt in similar ways to similar environments.
The best example of this are Marsupials. There exist, or have existed, marsupial equivalents to mammals, including squirrels, badgers, mice, wolves, sloths, herding animals, bears, and other common forms according to the ecology of Australia, which has ecological analogues to other parts of the world.
We can also see this in Batesian mimicry, the classic example of which is the viceroy butterfly, which has adapted to look like the monarch butterfly to deter predators. The monarch tastes bad, so predators will avoid it based on its distinctive wing pattern, which the viceroy has successfully mimicked.
There are many other examples of this.
Personally, I find it interesting that different kinds of trees are related more closely within their "plant family" than to other trees. Apple trees, for example, are in the same family as rose bushes and strawberry vines. Yet other trees, like redbuds and orchid trees, are in the same family as lupine bushes and legume vines like beans and peas. This despite the fact that the trees more resemble each other and operate more similarly than they do to their other family members.
6. Suboptimal design
There are, in some animals, what are called "Fixed Action Patterns" or FAPs, which are kind of like hard-wired software programs in their brains. One famous example of this is a cardinal that fed fish a typical cardinal chick diet over the course of several weeks. The cardinal's nest was destroyed and its chicks killed, but the cardinal, having the instinct for feeding, took care of the fish instead. Not optimal behavior to save the species.
A FAP found in human infants is the grasping reflex. Any mother with long hair will tell you what researchers found in a controlled environment: in the presence of hair, the hands of human babies grasp more strongly than with other materials. This FAP is also seen in primates and its purpose is for the young to hold on tightly to the hair of the mother as she moves quickly. This could be a vestigial behavior instinct, but I placed it here for continuity. There is no reason for a human baby to grasp hair more strongly than other textures.
You could also argue that the flounder's life cycle makes no sense. Aside from the eye moving from one side of the head to the other throughout the juvenile form, which is explainable, why doesn't the mouth alter to be flat as well? The mouths of adult flounder are arranged sideways as if the fish were still upright. Yet this does not offer an optimal form of feeding or other mouth behaviors as horizontal mouths do in other flat animals.
7. Geographical distributions
Australia's marsupials are a good example. It would be one thing if marsupials existed only in Australia, but the opossum exists in other parts of the world as well, pointing to the evolution of marsupials as a separate group, based on their geographic isolation after opossums appeared.
Similarly, monkeys are divided into "old world" -- Africa and Asia -- and "new world" -- the Americas. Both old and new world monkeys have opposable thumbs on their feet, but only old world monkeys have truly opposable thumbs on their hands. Nostril placement on the head, number of teeth, and prehensile tails are also differences between the two populations. There are also no "ground dwelling" new world monkeys.
8. Intermediate forms
Intermediate forms do not seem to be good examples of common descent because between any two intermediate form fossils is a perceived gap of an additional intermediate form. It therefore can be argued that all intermediate forms could have arisen at the same time in the same creation event for a purpose only known to the creator.

Post #49
This thread is interesting reading...I see a couple of issues that seem to come up often, both here and elsewhere. One is this:
For decades, clinicians have taken samples of influenza virus from individuals with the flu. In recent years, we have been able to determine the DNA sequence of the virus. We can, therefore, put together a diagram that shows the sequence relationships with respect to the actual timeline. Presumably, one of the viruses that was common in 1976 was the progenitor of some of the viruses that were common in 1977, and so on. By 1997, we have the descendents of that ancestral virus. Because influenza has a high rate of mutation, we see that the sequence keeps changing--but since we have the actual dates in which the samples were collected, we know exactly what order to arrange them in.
One such diagram is shown here. There are others around as well, but this is one that I could find easily, and that is accessible. Strains of virus that are very closely related (that differ by only a few bases) tend to be from similar locations in not-too-distant years. Strains that differ more significantly tend to be from more distant locations and more distant years. Still, by following sequence differences, we can put these different strains of virus together into a "family tree."
We really know a lot about this virus. The tree we draw fits with the known biology and genetics of the virus. BUT, you'll note that there are no individual virus strains at the nodes of the tree. Why not? Well, because we didn't happen to take a sample from the one individual who happened to have been infected with that particular virus that year. The fact that we don't have that particular sequence isn't a problem, though, because we know how influenza works. We know it is communicable. We know that, just like anything else, it reproduces according to its kind--so the virus from this year must be descended from the virus from last year. The link, or the "intermediate form" really existed, even if we didn't happen to sample it.
That is, we know that these different strains of virus are related by common descent from an earlier ancestor. The diagram simply shows us, in a convenient pictorial form, what the relationships are.
The particular tree that I've linked to here also shows another interesting thing. There is a cluster of related virus strains from Chile. There is another cluster of related strains from Europe. But, the Chilean strains are not so closely related to the European strains. Why? The simple answer is that Chile and Europe are far apart, so the Chilean virus and European virus had no interaction with each other. They were, if you will, separate populations of virus. In each population, mutations occurred at random--so they two populations picked up different mutations, and became different from their common ancestor and different from each other.
We see this trend much more markedly if we look at influenza in pigs, birds, horses, etc. Each has its own particular strains, each of which shows this kind of family tree. This is fundamental to understanding things like the 1918 pandemic, or the 1968 pandemic (which wasn't so bad), or the pandemic we're on the verge of having. Bird virus can't infect people, but both bird virus and human virus can infect pigs. In a co-infection in pigs, we can get recombinant virus that can infect humans, but escapes our immune systems because its antigenic proteins have evolved independently for years in the other host. We have no cross-immunity to it from prior infections, and we get really sick.
The point here is this: not only do DNA sequence comparisons give us trees that make sense (even if we don't have the sequences at the nodes), but we see that diversification into different varieties happens when populations are separated from each other.
These two principles apply to the much slower evolution of plants and animals (and other things). Common descent will give us these kinds of trees, but we probably won't be able to identify individuals at the nodes. Diversification will occur any time a population is split into two or more separate populations that can't interbreed (and therefore share genetic diversity). The nature of genetic inheritance (DNA is passed from parent to offspring), and the chemistry of DNA (occasional mutations occur) make it inevitable.
Since this much is, or should be, fairly straightforward (and is "only" microevolution), there shouldn't be any real difficulty with it. The big problem seems to come when we think about how speciation occurs. Can simple mutation and genetic divergence in two populations of the same species cause them to become different species? Well....
Here's a not-quite example. House finches were introduced into the US in the 1800's, on the east coast, and on the west coast. This established two populations. They have gradually migrated outward from their first locations--and are now meeting in the center of the US. The eastern population has distinct characteristics that make it distinguishable from the westerm population. They have become different. They are probably still able to mate with each other, and are therefore still the same species, but their morphological differences demonstrate the "proof of principle" that separate populations become different.
What would it take to make two populations like this not mate? With meadowlarks, the easter and western species sing different songs, and therefore don't recognize each other. Where their ranges overlap, they don't mate. So, here's one way to achieve reproductive separation. Another is to have the genes for the surface proteins of the egg and sperm "drift" in different directions, so that the sperm of one population can no longer interact effectively with the eggs from the other population. Each of these examples occurs through the normal, accepted processes of microevolution. The outcome--whether there is change in something "trivial" like feather color, or something "important" like successful mating--depends only on which genes are involved. Genes that govern song repertoire and recognition, or genes that govern sperm/egg interaction are just genes, and subject to mutation and genetic diversity, just like any others.
Once there is a barrier to mating between the two populations, they can't share genetic diversity ever again. They will continue to become more and more different, as they acquire different mutations and face different environmental selection pressures.
What happens if each of these two new species happens to split into a couple of separate populations? Each becomes different by the normal processes of microevolution.
In my opinion, the "hierarchical organization" issue boils down to DNA sequence comparisons. If two individuals are genetically related because they have a common ancestor, then they must share DNA sequences. The more closely related they are, the more DNA sequence information they will have in common. The less closely related they are, the less DNA sequence information they will have in common. When we compare a large number of living things, we find that there are DNA sequences that all of them have in common. We find that there are sequence differences as well, that allow us to build a tree of relationships. We don't have a time scale, as we do with the influenza tree, but we can still put the sequences together into a pattern of relationships.
Way out at the tips of the branches of the tree, there may be some variation in the DNA tree and the tree built from morphological characteristics, but overall, the general patterns of the DNA tree and the morphological tree are in remarkable agreement. This is pretty fascinating.
So, we are left with the following. The data give us a set of relationships, which we can present in a pictorial diagram as a tree. We may not have all of the intermediates on the tree, or examples at the nodes, but we have good examples that serve as "proof of principle." We know that descendents inherit their DNA from their ancestors. We know that DNA mutates, providing a source of genetic diversity. We understand the mechanisms, even if we don't have all of the examples for all species. The probability of creating this kind of genetic relationship among the earth's life forms by any mechanism other than common descent is vanishingly low. Unless, of course, it was Creation that was so designed as to give us exactly this series of relationships, down to the exact nucleotides in the DNA. Since we can't distinguish Creation with the Appearance of Common Descent from common descent itself, I'll accept common descent as the more plausible of the two.
In other words, the actual transitional form should exist and be identifiable. Well, let's consider the following:otseng wrote:If we are going to debate on the use of the hierarchical organization of life as evidence for common descent, I feel we should demonstrate exactly what we are referring to and use that as a basis of debate.
Also, in none of the diagrams I've seen, I have not seen any names of the animals/plants/organisms that is at or near the nodes of branching or splitting.
For decades, clinicians have taken samples of influenza virus from individuals with the flu. In recent years, we have been able to determine the DNA sequence of the virus. We can, therefore, put together a diagram that shows the sequence relationships with respect to the actual timeline. Presumably, one of the viruses that was common in 1976 was the progenitor of some of the viruses that were common in 1977, and so on. By 1997, we have the descendents of that ancestral virus. Because influenza has a high rate of mutation, we see that the sequence keeps changing--but since we have the actual dates in which the samples were collected, we know exactly what order to arrange them in.
One such diagram is shown here. There are others around as well, but this is one that I could find easily, and that is accessible. Strains of virus that are very closely related (that differ by only a few bases) tend to be from similar locations in not-too-distant years. Strains that differ more significantly tend to be from more distant locations and more distant years. Still, by following sequence differences, we can put these different strains of virus together into a "family tree."
We really know a lot about this virus. The tree we draw fits with the known biology and genetics of the virus. BUT, you'll note that there are no individual virus strains at the nodes of the tree. Why not? Well, because we didn't happen to take a sample from the one individual who happened to have been infected with that particular virus that year. The fact that we don't have that particular sequence isn't a problem, though, because we know how influenza works. We know it is communicable. We know that, just like anything else, it reproduces according to its kind--so the virus from this year must be descended from the virus from last year. The link, or the "intermediate form" really existed, even if we didn't happen to sample it.
That is, we know that these different strains of virus are related by common descent from an earlier ancestor. The diagram simply shows us, in a convenient pictorial form, what the relationships are.
The particular tree that I've linked to here also shows another interesting thing. There is a cluster of related virus strains from Chile. There is another cluster of related strains from Europe. But, the Chilean strains are not so closely related to the European strains. Why? The simple answer is that Chile and Europe are far apart, so the Chilean virus and European virus had no interaction with each other. They were, if you will, separate populations of virus. In each population, mutations occurred at random--so they two populations picked up different mutations, and became different from their common ancestor and different from each other.
We see this trend much more markedly if we look at influenza in pigs, birds, horses, etc. Each has its own particular strains, each of which shows this kind of family tree. This is fundamental to understanding things like the 1918 pandemic, or the 1968 pandemic (which wasn't so bad), or the pandemic we're on the verge of having. Bird virus can't infect people, but both bird virus and human virus can infect pigs. In a co-infection in pigs, we can get recombinant virus that can infect humans, but escapes our immune systems because its antigenic proteins have evolved independently for years in the other host. We have no cross-immunity to it from prior infections, and we get really sick.
The point here is this: not only do DNA sequence comparisons give us trees that make sense (even if we don't have the sequences at the nodes), but we see that diversification into different varieties happens when populations are separated from each other.
These two principles apply to the much slower evolution of plants and animals (and other things). Common descent will give us these kinds of trees, but we probably won't be able to identify individuals at the nodes. Diversification will occur any time a population is split into two or more separate populations that can't interbreed (and therefore share genetic diversity). The nature of genetic inheritance (DNA is passed from parent to offspring), and the chemistry of DNA (occasional mutations occur) make it inevitable.
Since this much is, or should be, fairly straightforward (and is "only" microevolution), there shouldn't be any real difficulty with it. The big problem seems to come when we think about how speciation occurs. Can simple mutation and genetic divergence in two populations of the same species cause them to become different species? Well....
Here's a not-quite example. House finches were introduced into the US in the 1800's, on the east coast, and on the west coast. This established two populations. They have gradually migrated outward from their first locations--and are now meeting in the center of the US. The eastern population has distinct characteristics that make it distinguishable from the westerm population. They have become different. They are probably still able to mate with each other, and are therefore still the same species, but their morphological differences demonstrate the "proof of principle" that separate populations become different.
What would it take to make two populations like this not mate? With meadowlarks, the easter and western species sing different songs, and therefore don't recognize each other. Where their ranges overlap, they don't mate. So, here's one way to achieve reproductive separation. Another is to have the genes for the surface proteins of the egg and sperm "drift" in different directions, so that the sperm of one population can no longer interact effectively with the eggs from the other population. Each of these examples occurs through the normal, accepted processes of microevolution. The outcome--whether there is change in something "trivial" like feather color, or something "important" like successful mating--depends only on which genes are involved. Genes that govern song repertoire and recognition, or genes that govern sperm/egg interaction are just genes, and subject to mutation and genetic diversity, just like any others.
Once there is a barrier to mating between the two populations, they can't share genetic diversity ever again. They will continue to become more and more different, as they acquire different mutations and face different environmental selection pressures.
What happens if each of these two new species happens to split into a couple of separate populations? Each becomes different by the normal processes of microevolution.
In my opinion, the "hierarchical organization" issue boils down to DNA sequence comparisons. If two individuals are genetically related because they have a common ancestor, then they must share DNA sequences. The more closely related they are, the more DNA sequence information they will have in common. The less closely related they are, the less DNA sequence information they will have in common. When we compare a large number of living things, we find that there are DNA sequences that all of them have in common. We find that there are sequence differences as well, that allow us to build a tree of relationships. We don't have a time scale, as we do with the influenza tree, but we can still put the sequences together into a pattern of relationships.
Way out at the tips of the branches of the tree, there may be some variation in the DNA tree and the tree built from morphological characteristics, but overall, the general patterns of the DNA tree and the morphological tree are in remarkable agreement. This is pretty fascinating.
So, we are left with the following. The data give us a set of relationships, which we can present in a pictorial diagram as a tree. We may not have all of the intermediates on the tree, or examples at the nodes, but we have good examples that serve as "proof of principle." We know that descendents inherit their DNA from their ancestors. We know that DNA mutates, providing a source of genetic diversity. We understand the mechanisms, even if we don't have all of the examples for all species. The probability of creating this kind of genetic relationship among the earth's life forms by any mechanism other than common descent is vanishingly low. Unless, of course, it was Creation that was so designed as to give us exactly this series of relationships, down to the exact nucleotides in the DNA. Since we can't distinguish Creation with the Appearance of Common Descent from common descent itself, I'll accept common descent as the more plausible of the two.
Post #50
Bump!
Endogenous retroviral sequences quite conclusively demonstrate common descent. Before i start, i'm not an expert on biology, so i'll have to simplify things in order to be able to talk about it
Retroviruses reproduce by inserting their own DNA into that of a cell, which then starts building more of those virii. Usually this is lethal to the cell, which at one time bursts.
However, occasionally it happens that the insertion of the virus' DNA is incorrect, and that it does not turn the cell into a virus factory. It survives the infection.
In even more rare cases it happens that this cell is a semen cell or an ovary (or one of those cells which are responsible for the production of those).
If this happens, then the individual that grows from it will have the inserted DNA in ALL its own cells, including the sex cells of that individual. This way the inserted sequence of DNA can spread through the population, by genetic drift, over the course of many generations (unless it is seriously detrimental, that is).
This has happened in the past, and such sequences could be identified in humans and other primates.
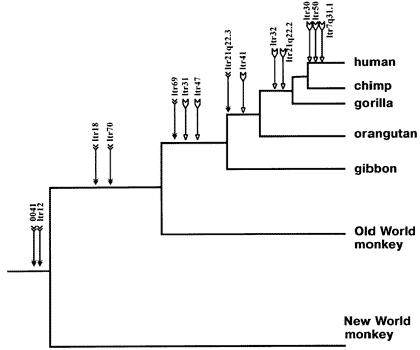
Humans have some unique sequences, but we also share some with chimps and gorillas. No other primate has this sequence in its genome, which is exactly what one would expect if the insertion happened before the speciation events that divided humans, chimps and gorillas from each other.
There are several other such sequences, and they fit exactly the phylogenetic tree of the theory of evolution. We do not see such sequences being shared by humans and orang utans, which gorillas and chimps do not have too.
This is a particularly good evidence for common descent, because the "common designer" argument doesn't work here. These sequences are not the result of design (or evolutionary development), but of an infection with a specific type of retrovirus.
Coincidence is extremely unlikely too, as the sequences do not only fit the predicted pattern of distribution, but they also are found in exactly the same part of the genome of the affected species - and retroviral insertions otherwise happen at a random place. One out of like six billion possible places each.
PS Even if we assume the chance that a sex cell is infected with the respective type of retrovirus and that the chance that the infection "fails" in the same way in all cases to be 1 (because we cannot really assign a probability for this), the odds of this happening by concidence without common descent are about 1: 10^460
jwu
Endogenous retroviral sequences quite conclusively demonstrate common descent. Before i start, i'm not an expert on biology, so i'll have to simplify things in order to be able to talk about it
Retroviruses reproduce by inserting their own DNA into that of a cell, which then starts building more of those virii. Usually this is lethal to the cell, which at one time bursts.
However, occasionally it happens that the insertion of the virus' DNA is incorrect, and that it does not turn the cell into a virus factory. It survives the infection.
In even more rare cases it happens that this cell is a semen cell or an ovary (or one of those cells which are responsible for the production of those).
If this happens, then the individual that grows from it will have the inserted DNA in ALL its own cells, including the sex cells of that individual. This way the inserted sequence of DNA can spread through the population, by genetic drift, over the course of many generations (unless it is seriously detrimental, that is).
This has happened in the past, and such sequences could be identified in humans and other primates.
Humans have some unique sequences, but we also share some with chimps and gorillas. No other primate has this sequence in its genome, which is exactly what one would expect if the insertion happened before the speciation events that divided humans, chimps and gorillas from each other.
There are several other such sequences, and they fit exactly the phylogenetic tree of the theory of evolution. We do not see such sequences being shared by humans and orang utans, which gorillas and chimps do not have too.
This is a particularly good evidence for common descent, because the "common designer" argument doesn't work here. These sequences are not the result of design (or evolutionary development), but of an infection with a specific type of retrovirus.
Coincidence is extremely unlikely too, as the sequences do not only fit the predicted pattern of distribution, but they also are found in exactly the same part of the genome of the affected species - and retroviral insertions otherwise happen at a random place. One out of like six billion possible places each.
PS Even if we assume the chance that a sex cell is infected with the respective type of retrovirus and that the chance that the infection "fails" in the same way in all cases to be 1 (because we cannot really assign a probability for this), the odds of this happening by concidence without common descent are about 1: 10^460
jwu
Last edited by jwu on Fri Dec 24, 2004 7:19 am, edited 2 times in total.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}