
Did humans descend from other primates?otseng wrote: Man did not descend from the primates.
Are humans primates or should there be special biological taxonomy for humanity?
Please cite evidence.
Moderator: Moderators

Did humans descend from other primates?otseng wrote: Man did not descend from the primates.
Well you're wrong on the 'semi-pedal' comment straight off the bat given that almost all primates can move biped-ally to some degree. Some are very good at it, and others not.otseng wrote:I'll quote Juan Luis Arsuaga:McCulloch wrote: What is it about humans that you believe is novel, as compared to the other primates?
The Neanderthal's Necklace: In Search of the First ThinkersWe are unique and alone now in the world. There is no other animal species that truly resembles our own. A physical and mental chasm separates us from all other living creatures. There is no other bipedal mammal. No other mammal controls and uses fire, writes books, travels in space, paints portraits, or prays. This is not a question of degree. It is all or nothing; there is no semi-pedal animal, none that makes only small fires, writes only short sentences, builds only rudimentary spaceships, draws just a little bit, or prays just occasionally.
McCulloch wrote: What is it about humans that you believe is novel, as compared to the other primates?
OK, and I'll quote the linked review of the same bookotseng wrote: I'll quote Juan Luis Arsuaga:
The Neanderthal's Necklace: In Search of the First ThinkersWe are unique and alone now in the world. There is no other animal species that truly resembles our own. A physical and mental chasm separates us from all other living creatures. There is no other bipedal mammal. No other mammal controls and uses fire, writes books, travels in space, paints portraits, or prays. This is not a question of degree. It is all or nothing; there is no semi-pedal animal, none that makes only small fires, writes only short sentences, builds only rudimentary spaceships, draws just a little bit, or prays just occasionally.
[font=Times New Roman]emphasis mine[/font]The Neanderthals provide a surprising mirror for modern-day humanity. They belonged to our evolutionary group and lived like the Cro-Magnons, our ancestors, did " worshipping, socializing, and hunting. The struggle between Neanderthals and Cro-Magnons lasted thousands of years. The Cro-Magnons were not biologically fit for extreme cold weather, but their ingenuity allowed them to settle down, band together, and survive. In this tale of life, death, and the awakening of human awareness, Juan Luis Arsuaga, Spain's most celebrated paleoanthropologist, depicts the dramatic struggle between two clashing species, of which only one survives.
Bonobosthere is no semi-pedal animal

elephantsdraws just a little bit
Again, bonobos(but there are several other examples in chimps and orangutanswrites only short sentences
Neither did any man do so until the 1940s, this is a ridiculous statement. But a space ship is a tool...builds only rudimentary spaceships,
"In an outing in the Georgia woods, Kanzi touched the symbols for "marshmallows" and "fire." Susan Savage-Rumbaugh said in an interview that, "Given matches and marshmallows, Kanzi snapped twigs for a fire, lit them with the matches and toasted the marshmallows on a stick.""none that makes only small fires
Not sure how Juan read the minds of any creature. And I'm not sure how praying is different in kind from cowering in fear of anything a creature does not understand.or prays.

Trivially, any process that takes longer than a human lifetime (or the lifetime of human civilization) is unobservable. However, this is a different than saying that the mechanisms which underlie the process are unobservable.otseng wrote: There's nothing wrong with extrapolation by itself. To reiterate my point, all I'm saying is that macroevolution is unobservable. I brought this up originally to counter the charge that a designer should be dismissed because it cannot be observed and revealing the inconsistency in the use of observability.
You've provided an example spanning a few millennia. The timespan in question is millions of years. Why should we expect to see the same degree (quantitative) of genotypic change (and corresponding phenotypic change) on human timescales? Again, unless you will describe a difference of mechanism for macroevolution (i.e., why you have reason to suspect observed mechanisms are incapable of accounting for the diversity among species), you aren't making much of a case against the validity of the evolutionary extrapolation. The whole purpose of this exercise is to extrapolate beyond historic timescales (many times the scope of all recorded human experience, which goes back only some thousands of years).What I have shown is that from human experience in the domestication of animals, there is not much significant change in morphological features in animals to account for common descent.
'Species' is a useful but arbitrary label. I maintain that there is no such thing as an ideal horse or ideal oak tree that exists independently of our minds - these are only labels we apply to biological replicator populations with similar genomes (and even the genetic contents of those populations change over time and generations). I'm all for the replacement of Linnaean taxonomy with something like Phylocode. But that's neither here nor there.Yes, I would agree that there is a subjective element to it. But even the term "species" is a bit subjective.
...
And since micro and macro is often differentiated at the species level by evolutionists, then the terms microevolution and macroevolution are also likewise subjective.
I'm not so sure there can be an objective evaluation [of macro- vs. microevolutionary traits] either. But, this would be a problem across the board.
This seems like a dead-end argument. To the extent that a bee wing and a penguin wing are both flat, digitless structures protruding from the dorsal side of the organism (i.e., wings), theyre morphologically similar. We could argue similarity all day. Does your model provide any hard predictions about patterns of similarity in nature (either genetic or morphological)? Correspondingly, do these model predictions provide falsification criteria (patterns which falsify design if observed)?I think the difference here is "similarity of function" and "morphological similarities". Two different morphological features could be used for a similar function. A hoof and a webbed foot can be used for walking, but are not similar morphologically.
As I commented above, I think the best anyone can do is appeal to parsimony for model discrimination. We see a pattern in existing species and evolutionary biology provides a parsimonious and falsifiable explanation for that pattern based on observed mechanisms. Simply positing a designer does not make a designer more parsimonious (because it has not yet been observed if its even observable). Thus far you have been reluctant to discuss the designers tools (or we can call them mechanisms if you dislike tools). Trivially, if one posits a designer that created all life such that it appears the way it appears, it is unfalsifiable (besides being tautological). This statement also has zero explanatory power. If somebody presented it to you, why would you even consider it?otseng wrote:I try not to use that as an explanation and I don't believe I've ever stated that here in this thread.GrumpyMrGruff wrote:I can see no a priori reason to expect this pattern if the organisms were designed. Can you provide one? 'The designer could've done it that way' doesn't cut it. The designer could've done anything - you refuse to discuss the scope and limitations of its methods. What reasons do we have to think it should've done it that way?
Granting for the sake of argument that a viable evolutionary trajectory between chimps and humans could be found, it would not falsify claims of special creation for the reason given above. A dedicated creationist would have no reason to accept that, simply because a viable path exists, that path was ever actually traversed in the past. Certainly, such a finding would strengthen the evolutionary explanation (reinforcing the view that known mechanisms are sufficient to account for known biodiversity). Compare: If a creationist identified an Ice Age land-bridge between Asia and Australia, it wouldnt obligate non-Christians to accept the additional claim that marsupials ever traversed it on their way from Noahs Ark to Australia.The only way I can currently think of to falsify what I claim using molecular genetic evidence is what I stated earlier - "Genetic changes from one species to another and leading to humans are identified." For the purposes of this discussion, just the genetic steps from the common primate ancestor to humans would suffice to falsify the Human Creation Model. I would agree that modern genetics is not able to do this now. But, if it's ever done in the future, the model I proposed would be falsified.
I disagree. Both models need to be evaluated on their own merits to rule out the possibility that either of the models is actually unfalsifiable. Ive shown how the mechanistic predictions of evolutionary theory provide a falsification criterion " patterns of genetic data incompatible with the theory. You have yet to show that your model is even capable of providing similar predictions and falsification criteria. (And if it cant, why are we considering an unfalsifiable model?)I think this would apply if there was a third explanation on the table. If there are only two, then it is fair to compare and contrast the two.
Again, if you dislike tools, how about mechanisms? Im not talking about knowing the brand name of jackhammer used to carve Teddys nose, but rather the assortment of mechanisms available to fashion the sculpture. Lets extend your Rushmore example. Mount Rushmore is suddenly transported to a mountain range on a distant planet. It is the metaphorical watch on Paleys now-cosmic beach. An alien flies by. It has never seen humans (or human faces) " heck, it doesnt even have eyes. How can it determine that Rushmore is artificial? Minimally, it needs to know the potential mechanisms used to construct it (contra the natural processes of the planet that mayve formed it). If you disagree, would you please explain?I was specifically addressing your point "You have been invoking a designer, but you have not specified any of the tools used by the designer (or the genetic artifacts left by those tools which we might observe today)." My point is not about the ability to detect design, but that knowledge of the tools used is not necessary.
If [identification of all intermediate genomes between species] is even impossible in principle, then there'd be no way to unequivocally prove common descent through genetic evidence.
This is incorrect. Phylogeny constructs trees based on the nested hierarch of genetic similarity in compared sequences. Mutation, drift, and natural selection provide a parsimonious explanation for the recurrence of the same tree when sampling across different sequences (design provides no such a priori mechanistic explanation for this observation). Some phylogenetic software attempts to reconstruct ancestral sequences at internal nodes of trees, but this step is not required for the process. At any rate, the programs have no rigorous way to deal with insertions and deletions. Consequently, they have no way of reconstructing whole genomes because whole genomes contain many variable insertion/deletions as a result of things like retrotranposons and transposons (among other mechanisms).If this is true (and if it's impossible to know the genome of extinct organisms), then it'd be impossible to construct a tree of life diagram based on genetics.
Id appreciate if you'd point out where youve offered definitive proof, because such proof is impossible within a scientific framework. Do you mean conclusive evidence? If so, where have you offered this? The best we can strive for is a predictive, therefore falsifiable (but-not-yet-falsified), parsimonious explanation that accounts for available data.What I've offered is the definitive proof and falsification of the theories. Can you offer then any other suggestions that would be the definitive proof and falsification of the theories?
Youre being a bit careless with proof again. I can see this going back and forth for a long time. Do you agree that mechanistic, falsifiable, parsimonious explanations are the best we can do? If so, I think Ive already made my case that evolution provides the most parsimonious explanation. Im waiting for you to make yours.But, if you say that [reconstructing the extinct genomes from known genomes] would be impossible even in principle in the future, then I'll defer to you to provide methods to prove common descent.
My purpose with this figure was to show that you are demanding too high a resolution from necessarily incomplete data.One might see [competing hominid phylogenies] as evidence of human evolution. I see it as evidence of the intractability of being able to coming to a consensus of arranging hominid evolution to man.

I think stating that there's "quite a bit of evidence" is overstating your case.SailingCyclops wrote:There's quite a bit of evidence:otseng wrote: Please show evidence of fire being in use in the last million years.
Tools older than 1.7 million years ago would be Mode One tools, which consist of stone pebble tools. If flint tool artifacts were found older than this, it would not fit into the normal tool evolutionary timeframe.Evolution of FireThe use of fire has long been thought to have coincided with, and perhaps aided in the precipitation of, the evolution of modern human culture and language around forty thousand years ago. That is, up until last year, when two controversial studies were released that brought this long held notion into question. These new findings suggest that not only might fire have been put to intentional use by humans as far back as 1.6 million years ...
[...]
Last April, this question was answered by Brian Ludwig of Rutgers University.
After studying forty thousand pieces of flint tool artifacts, ranging from 1 to 2.5 million years old, from sites throughout Africa, Ludwig found some surprising results. When rocks have been exposed to heat, they develop telltale signs of heat exposure, in the form of observable potlid fractures. After studying his tool artifacts, Ludwig discovered that the artifacts he was studying started to show these small fractures only after about 1.6 million years (McCrone, 2000). This finding suggests that by that time, H. erectus was not only using campfires regularly, but also hunting and possibly even cooking tools.
[...]
Yes, spotty though it may be.Human evolution, fire, and food... In spite of the fact fire use becomes spottier the farther back in the historical record you go, Wringham maintains that this clutch tool could have triggered the cascade. In fact, it is interesting to note that evidence for the use of fire "spotty though it may be- goes as far back as 1.6 millions years to about the time H.erectus appeared.
[...]
Another interesting quote from your source:Did homo erectus discover fire?The homo sapiens mental revolution took place between 100,000 and 40,000 years ago, following the development of grammatical speech. But Homo erectus was pretty smart already judging by evidence that fire had been discovered over 1.6 million years ago. Could fire really have been such an early discovery?
[...]
New evidence, however, suggests that human exploitation of fire may be quite incredibly ancient, going back some 1.6 million years. Recently developed forensic techniques are strengthening the case that some long-disputed fire remains found in Kenya, East Africa, were indeed kindled by our ancestors.
[...]
At both sites, archaeologists found the bones and stone tools of Homo erectus--the first hominid species to have a markedly larger brain and fully human-proportioned body. At Koobi Fora, the excavations also uncovered a scattering of ten small, half-metre diameter, "lenses" of baked orange earth dating to around 1.6 million years ago.
[...]
Within palaeoanthropological circles, there will be many fingers crossed hoping the latest findings just arent true. This is because hominid control of fire at 1.6 million years poses huge problems for current thinking about human evolution. The story goes that technologically sophisticated humans arrived with a "big bang" only about 40 000 years ago, with the development of grammatical speech. If the very early date for the control of fire holds up, then either we have to believe that kindling a roaring blaze is essentially--indeed literally--a pretty dumb skill. Or else we must be willing to upgrade the mental abilities of our forebears rather considerably.
As Zeeby pointed out, in stochastic birth-death models like this the number of progeny left by each member of the initial population will inevitably go to zero or one after a sufficient amount of time (dependent on model parameters). The correct question is not if one female's offspring will eventually dominate the population, but how long (on average) it will take for this to occur. To answer that question, we need to know things like population structure, generation time, population size, etc.otseng wrote:However, the question is what can explain that all other female lines disappeared? In terms of the probability that only one female is the progenitor of all, I'm going to ask Zeeby in my next post about that.
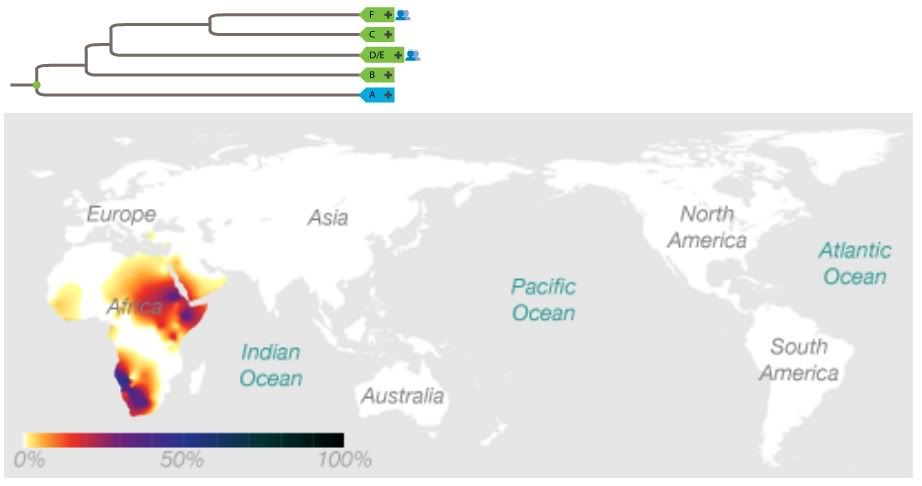
True. You know what they say about sailors: A girl in every port. Mediterranean, coastal Atlantic, and over-land trade (including Mideast and African slave trade) dates back thousands of years, at least to the Phoenicians. There was doubtless some gene flow between these populations. However, the majority of living haplogroup A members live in Africa (all figures adapted from 23andme.com unless otherwise noted):Haplogroup A is not limited to Africa.
http://en.wikipedia.org/wiki/Haplogroup_A_%28Y-DNA%29Haplogroup A is found mainly in the Southern Nile region and Southern Africa. However, at lower frequencies, M91 is found in many areas of Africa, including Morocco, Egypt, and Cameroon. Outside of Africa, it has been detected in European males in England, Portugal, the Mediterranean islands of Sardinia, Italy, and Lesbos, Greece, and the Eastern Mediterranean regions of Anatolia, the Levant, and Southern Arabia.
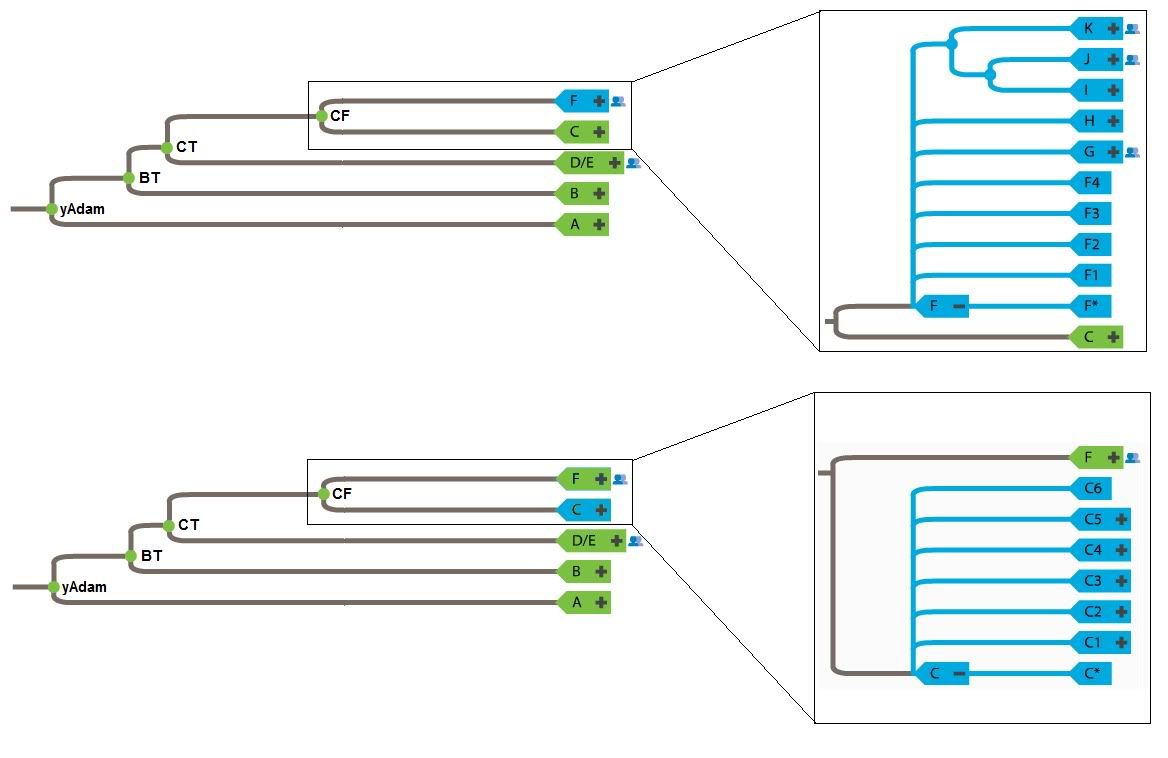
It would be correct to say that nobody sampled has the BT*, CT*, or CF* haplotype. Conversely, greater than 90% of the world has the mutational markers that denote the BT haplogroup. What does this mean? The asterisk denotes an ancestral state. Consider the following examples.For Haplogroup BT, it is a hypothetical grouping and no human actually has a BT Y-chromosome. Same also goes for CT and CF.
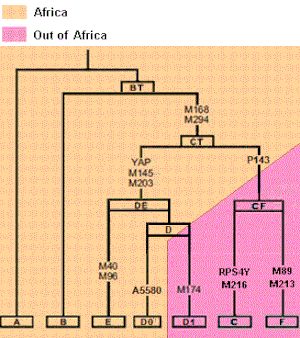
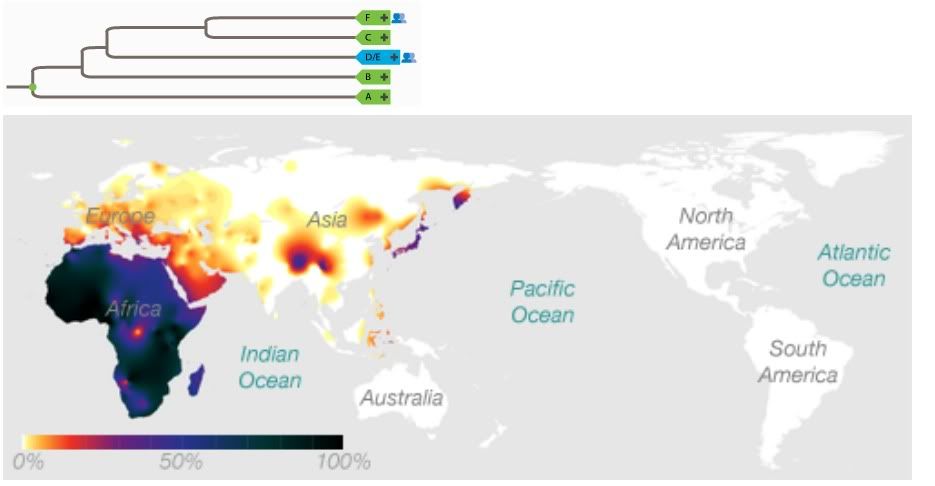
Yet if you map majority biogeography onto the phylogeny (as shown in the wiki page you link to), an interesting trend emerges.Also there is no definitive place of origin for BT either, though sources say that it probably originated in North East Africa.
http://en.wikipedia.org/wiki/Haplogroup_BT_%28Y-DNA%29Haplogroup BT split off from haplogroup A 70,000 years bp , probably originating in North East Africa from Y-chromosomal Adam. It contains all living human Y-DNA haplogroups except for A.
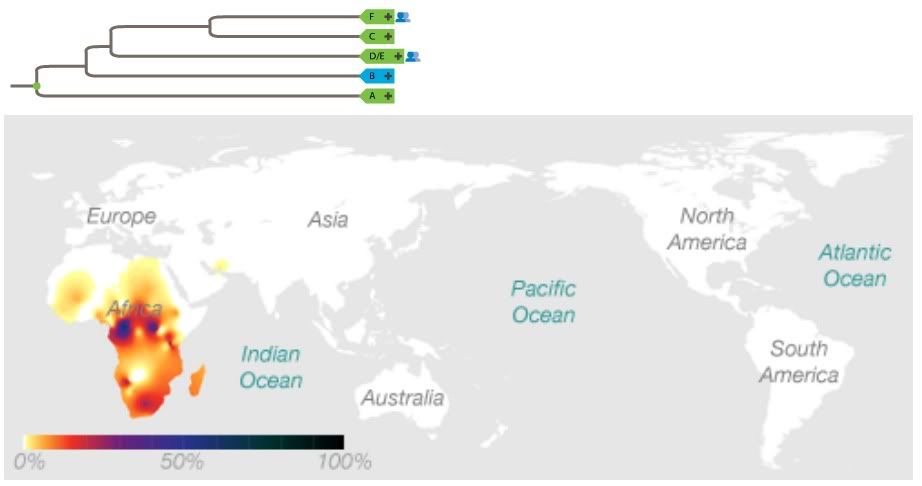
Yep, most Eurasians are. I'm a member of F. Specifically, G2a. I don't feel special at all.The F Haplogroup contains more than 90% of the world's population.
http://en.wikipedia.org/wiki/Haplogroup_F_%28Y-DNA%29In human genetics, haplogroup F or FT is an enormous Y-chromosome haplogroup spanning all the continents. This haplogroup and its subclades contain more than 90% of the world's existing male population.
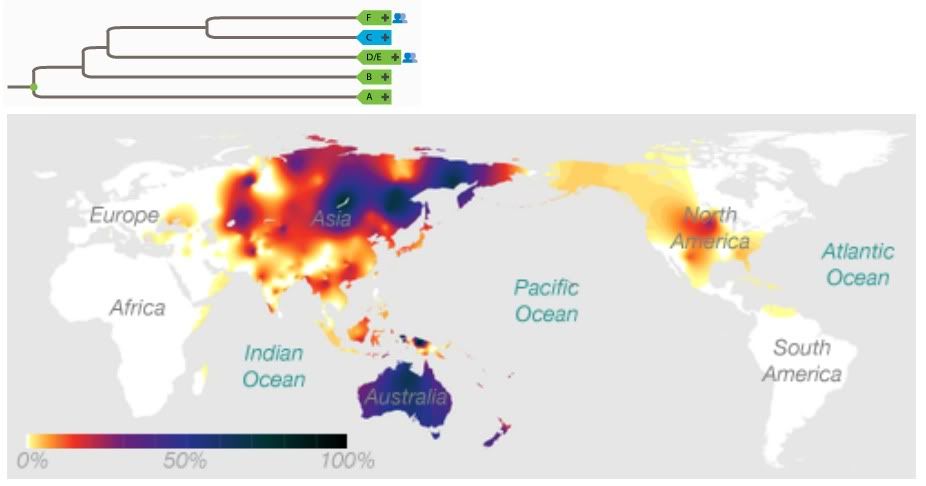
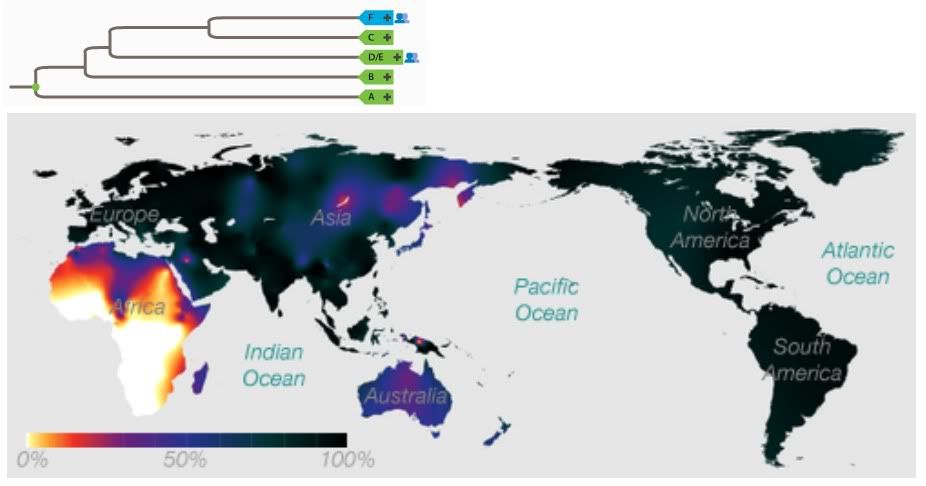
Well, somewhere ranging between the Mideast and India, anyway.It is theorized that it originated in the Middle East area.
You lost me here. This sounds like a non sequitur. yAdam is hypothetical in exactly the same way that BT, CT, and CF are hypothetical: Their markers, though shared by many males (the default marker set in the case of yAdam) are not found in isolation in any living male.Because BT, CT, and CF are all hypothetical and not found in any human, it is entirely possible that F can be located nearer to the root of the tree.
This statement ignores the important fact that we cannot assume identical birth and mortality rates for all populations (dependent on environment and lifestyle). This might be a valid extrapolation if we assumed lockstep reproduction for all humans and ignored both phylogenetic and biogeographical evidence.And since it accounts for 90% of the current population, this would make it seem more likely that it is close to the root.
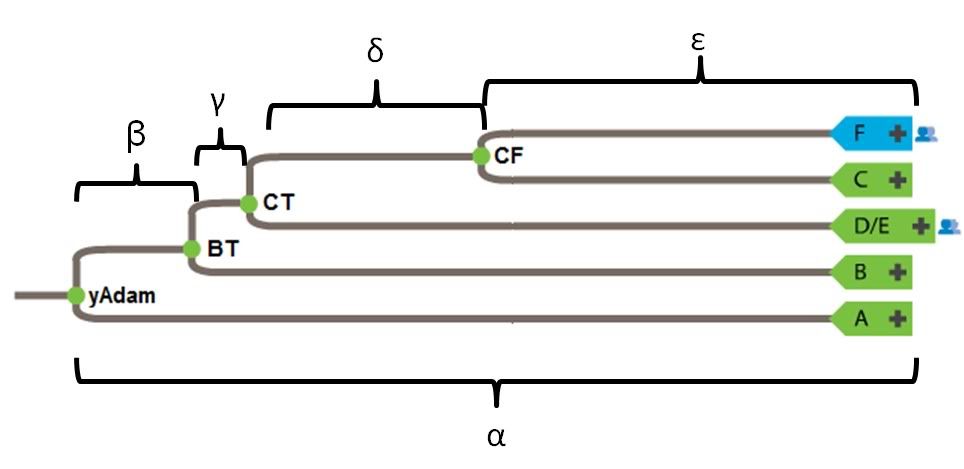
Upon inspection this tree presents a major problem. Phylogenetic analysis suggests that B is more closely related to D/E, C, and F than it is to A. Your creation of an AB most recent common ancestor is at odds with the genetic data. Aside from being wrong (insofar as it contradicts data), the root of this tree could be placed in either Africa or Eurasia. It doesn't do much to advance your case for a Eurasian origin.So, though this is tentative for now, it's possible to arrange the Y-chromosome tree into 3 descendants off of a common ancestor: AB, CF, and DE.
This is why fossil evidence represents necessarily insufficient data set for unique phylogeny determination, yes.Then the use of fossil evidence would be unreliable.GrumpyMrGruff wrote: If the only features we could compare were morphological, there might be cause for confusion (since different sequences can sometimes encode similar structures).
Yes. There are different methods, but in general they arrange the sequences so that same mutations don't have to occur over and over again (an unlikely event). In other words, they assign similar sequences a more recent common ancestor than dissimilar sequences because their similarity is most parsimoniously accounted for by an ancestor that possessed their shared traits. As I've said, this is not the interesting part. The interesting part is that new sets sequences from from same set of organisms yield the same tree when you apply this method. This is what's predicted in new data if mutation and natural selection are driving the process, but this pattern is not accounted for a priori by design models. Creationists must apply a post hoc, non-mechanistic explanation that 'the designer could've built them this way' (unfalsifiable). How couldn't the designer have done it?Would even phylogenetic trees based on genetics also assume homology?GrumpyMrGruff wrote:Note that every phylogenetic tree we make assumes homology.
I am waiting on a phylogenetics textbook I ordered this week. Until it arrives, I will only be able to proceed off of class notes from a phylogenetics course I took a few years ago. With your permission, I'd like to get my references together before I go into this in detail.OK, let's spend some more time on this then.GrumpyMrGruff wrote:simply show that different genes from the same set of organisms infer the same tree no more often than randomized trees.
If different genes from the same organisms point to different evolutionary pathways, these would be different trees correct?
What would constitute randomized trees?
Ultimately, the scientific method is used to ask and answer "How?" Invoking design does not answer how a designer did anything, and we cannot make falsifiable predictions about new data without determining what physical characteristics arise from the design process. Remember, predictions come in the form: 'New data I acquire should have the patterns ABC if they were caused by Mechanisms XYZ.' Falsification criteria come in the form: 'Mechanisms XYZ are incosistent and/or insufficient to cause the patterns DEF; if we see DEF, XYZ are wrong/insufficient.'In terms of the mechanism that a species is designed, it not necessary to know how something is designed to posit a designer.
On the contrary, SETI made strong assumptions about the mechanisms used by ETIs: They assumed any intelligence-based broadcasts would be produced by frequency or amplitude modulation of EM waves (radio, microwave, etc.). These are the types of signals they listened for. If ETIs exist and use other methods of encoding information, SETI (because of its assumptions about how ETIs created their signals) would have missed them.One example would be the SETI program. If researchers find a signal from another planet, it would not be immediately rejected because they do not know how an alien civilization created the signal.
Unless of course the ETI was intentionally transmitting a signal intended to be understood by other technological civilizations as originating from intelligence. Such a transmission could be as simple as a set of prime numbers being sent as frequency modulated tones. Receiving such would certainly rule out natural causes. I know of no quasar which could mimic a prime number em burst.GrumpyMrGruff wrote:Sometimes a natural phenomenon provides a more parsimonious answer, but the only way to confirm ETI sources for signals would be to attempt back-and-forth communication (likely impossible due to distance and conceptual barriers) or attempt to sample the source directly (likely impossible due to distance).
As a sci-fi nerd, I love the idea of SETI. As a scientist, I think it was a questionable investment at best. It attempts to address a question that seems practically unfalsifiable in most cases (even if it is falsifiable in principle).
I agree that we would flag a prime number signal as potential ETI because a) we know of no astronomical phenomena that produce such transmissions and b) we can produce them ourselves. My contention is that having intercepted such a signal, there are no practical ways to verify or rule out an ETI source (with a few exceptions if the source is exceptionally nearby).Unless of course the ETI was intentionally transmitting a signal intended to be understood by other technological civilizations as originating from intelligence. Such a transmission could be as simple as a set of prime numbers being sent as frequency modulated tones. Receiving such would certainly rule out natural causes. I know of no quasar which could mimic a prime number em burst.
Bob



{kind=link}